易基因综述:表观遗传学和表观育种在作物品种改良的重要作用(水稻+玉米+番茄+大豆+油菜)
大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
2022年8月15日,《Front Plant Sci》杂志发表了题为“Epigenetics and its role in effecting agronomical traits”的综述文章,讨论了DNA甲基化、组蛋白修饰、RNA修饰等表观遗传学在影响水稻、玉米、番茄、大豆、油菜等作物性状中的作用。

期刊:Front Plant Sci
日期:2022.08.15
IF:6.627 /Q2
摘要
为养活不断增长的人口,迫切需要具有适应气候变化能力的作物。因此开发具有较好农艺性状的高产作物品种是农业研究中的重要问题。大多农艺性状是定量的,且受到复杂的遗传调控阻碍作物改良。植物表观育种是一种利用表观遗传变异进行作物生长发育的方法,在作物改良领域具有广泛应用。表观遗传学是指通过DNA甲基化、组蛋白翻译后修饰、RNA干扰诱导的可遗传基因表达变化,表观遗传修饰通过改变染色质状态来影响基因表达,染色质状态促进植物生长并决定对外源和内源环境的表型响应。表观遗传修饰可以通过提供有用的标记来改良育种,同时还考虑表观基因组多样性以预测植物表现并提高作物产量。本综述重点总结了表观遗传变化在表观育种中对作物改良的作用。着眼于表观遗传变化在作物品种改良中的重要性,全面总结了表观遗传分子方面的前沿知识及其在作物育种中的应用。
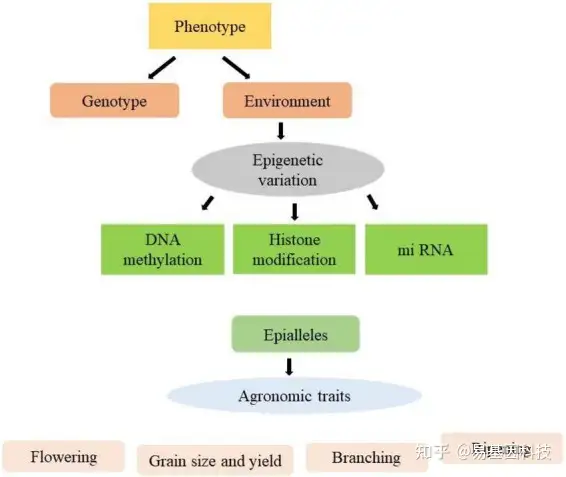
图1:利用表观等位基因(epialleles)改良各种农艺性状
植物表观育种
传统作物育种包括杂交、选择所需性状的遗传变异,导致遗传基础缩窄和遗传多样性缺失,从而阻碍作物改良。表型性状受遗传学和表观遗传学影响,利用表观遗传变异或表观基因组变化可以提高作物对环境条件变化的响应,保证农业产量和质量。先前研究已经鉴定出许多调控所需性状的基因或QTL(quantitative trait locus),但遗传力缺失仍然是影响表型的关键障碍。
表观遗传变化是遗传力缺失主要原因之一,表观等位基因是具有表观遗传修饰的位点,可以自然产生或人工诱导稳定传递到子代。除促进表型多样性的自然遗传变异外,表观等位基因还可以提供遗传变异的其他来源(图2)。表观等位基因主要包括表观遗传(不遗传)和遗传两种形式。不遗传性表观等位基因来源于导致染色质状态变化的发育因子或环境信号,也可由自发的表观突变引起,如拟南芥胞嘧啶的自发表观突变率比基因突变率高1000倍。大豆、玉米和番茄等作物中多胞嘧啶位点的差异甲基化区域(DMR)不太常见。此外,由于在基因间区域插入转座子而产生遗传性表观基因,通过DNA甲基化而导致其失活。在拟南芥、水稻和玉米中有过报道,DNA甲基化增强位点两侧甲基化,导致新的表观基因形成。
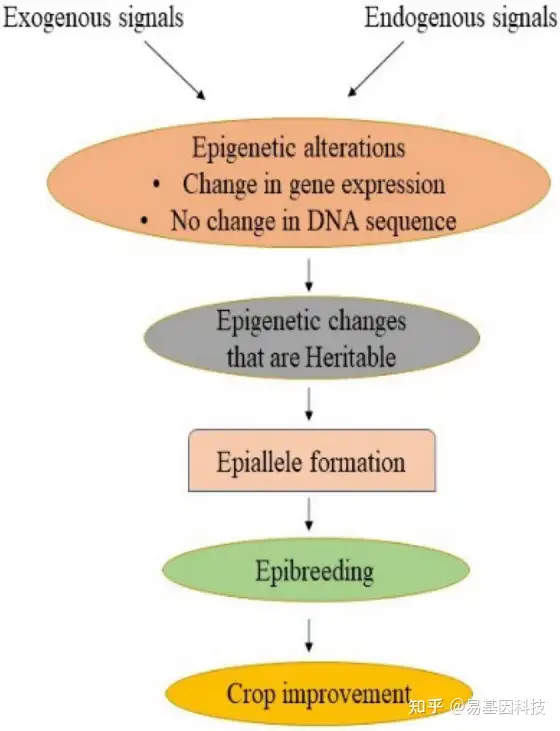
图2:表观遗传修饰可以响应外部和内部环境,导致基因表达变化而不改变DNA序列。稳定和可遗传的表观遗传变化导致表观等位基因的形成,可用于作物改良的表观育种计划。
表观遗传学对不同作物农艺性状的影响
水稻
水稻(Oryza sativa L.)是一种重要的谷物作物,作为主食养活了世界一半以上的人口。表观等位基因epid1编码GTP结合蛋白会导致生长发育迟缓。d1基因通过DNA甲基化和组蛋白乙酰化沉默,导致水稻植株高度调节。OsSPL14是一个与富民基因(WEALTHY FARMERS PANICLE,WFP)相关的表观等位基因,表观遗传变化增加OsSPL14基因表达,导致稻穗分枝和籽粒产量增加。
开花期(抽穗)是水稻籽粒形成的最重要阶段,受表观遗传修饰调控,早花导致产量下降,而延迟开花导致结实率下降。组蛋白甲基转移酶(HMTase)基因,如SET结构域基因724(SDG724)作为SET域家族II类,通过甲基化组蛋白H3赖氨酸36(H3K36)促进开花。在长日照(LD)和短日照(SD)条件下,SDG724功能缺失突变体lvp1均表现出开花延迟,这与水稻开花位点T1(RFT1)表达下调以及抽穗期3a(Hd3a)有关。尽管这两个基因长度约11.5 kb,但只有RFT1的染色体区域通过H3K36me2/3修饰降低了转录活性。在另一项研究中,在转录起始位点周围H3K9乙酰化,可以增强Hd3a RNAi转基因植物中RFT1表达。SDG725也属于SET域家族II类,通过H3K36me2/3促进开花。OsTrx1属于SET结构域家族中III类,可以活化或维持转录基因的活性状态,还可以延长LD植物的开花期。WOX11(Wuschel相关同源-box基因)通过ADA2-GCN5组蛋白乙酰转移酶调控与冠根发育相关基因。QTL OsglHA T1(一种新型GNAT样蛋白)具有内置的H4组蛋白乙酰转移酶活性,并与粒重相关。PGL2(6-PHOSPHOGLUCONOLACTONASE 2)由OsglHAT1正调控,对粒长至关重要。OsglHA T1表达上调对粒长、粒重、产量和总生物量等农艺性状均有积极影响。
在水稻中,OsSRT1是一种SIR2型HDAC,可抑制糖酵解通路的碳代谢通量,同时增强生长种子中淀粉积累。叶角是直接影响植物结构和谷物产量的另一个农艺特征,直立叶片的植物可以吸收更多阳光进行光合作用,增加籽粒灌浆的氮储存,从而增加产量,且适合密集种植。叶角、粒径和产量潜力受油菜素类固醇(BR)植物激素调控,当BR生物合成基因过表达时,叶片变得不直立,且具有相当大的叶倾斜;而在BR缺失或BR不敏感突变中,叶片直立。由于RAV6启动子区低甲基化,表观等位基因Epi-rav6与较大的叶片倾斜度和较小的晶粒大小有关。编码转录因子家族(TCP OsPCF7)基因在植物结构中起作用,OsPCF7在转基因水稻幼苗中表达增加,提高了枝条高度、根长、根数、分蘖和抽穗率,从而导致单株籽粒产量增加。启动子DNA甲基化可以沉默营养组织中的OsFIE1,异位OsFIE1表达由启动子DNA甲基化缺失的表观突变引起,导致数百个基因中的矮化、花异常和H3K27me3水平变化。ESP基因低甲基化与穗结构调控有关,这些植物表现出短而密的穗。SE1缺失或阻遏复合组分功能受损阻碍了Eui1区域组蛋白去乙酰化和H3K27me3甲基化,导致染色质从闭合状态转为开放状态,从而增加Eui1转录,降低赤霉素并导致矮化表型。NGR5通过促进PRC2(Polycomb repressive complex 2)招募以通过H3K27me3(组蛋白H3赖氨酸27三甲基化)限制芽分枝来抑制基因表达,从而促进分蘖以响应氮供应的增加。启动子区域OsAK1基因高甲基化与光合能力相关,OsSPL14基因受microRNA调控,并影响穗分枝和产量增加。miR160通过下调OsARF18表达影响生长素信号传导,从而影响水稻生长发育。miR172靶向AP2 TF并影响花发育、开花时间和穗分枝。通过osa-miR397产生漆酶样蛋白下调OsLAC基因,可以调控粒径和产量。
玉米
玉米(Zea mays L.)是全世界都在种植的重要主食作物。2012年,Barbara McClintock在玉米中发现了转座子沉默的表观遗传过程,通过转座因子(TE)减少转座和基因组破坏。活性TE通常与低甲基化相关,而TE沉默显示出显著水平的DNA甲基化。植物的许多过程,包括发育调控、外部刺激响应和适应,都受到通过DNA/RNA甲基化、翻译后组蛋白修饰和ncRNA的表观遗传调控影响。ZmMRP4基因甲基化产生lpa1-241表型,种子中含有大量无机磷酸盐。另一项研究中,p1基因甲基化(Myb同源并调控红色素生物合成)与色素沉着减少有关。HDA108作为一种组蛋白去乙酰化酶,通过减少H3K9二甲基化和增加细胞核中的H3和H4组蛋白乙酰化来调控植物高度、叶片发育和繁殖力。HDA101组蛋白去乙酰化酶基因下调和过表达会影响乙酰化组蛋白数量,导致形态和发育缺陷。HDA101与共抑制剂(如NFC103/MSI1和SNL1/SIN3样蛋白)互作导致靶基因上组蛋白H4赖氨酸5(H4K5ac)低乙酰化,但不影响其转录水平,其通过调控细胞特异性基因表达转移来影响内核大小,切除会导致目标区域组蛋白高乙酰化而不影响转录水平。此外,HDA101也针对一些与高乙酰化和增强表达相关的非活性基因。
ZmGCN5通过与衔接蛋白ZmADA2和bZIP转录因子ZmO2互作,促进种子成熟过程的胚乳发育。DNA甲基转移酶基因(dmt102和dmt103)通过DNA去甲基化沉默,导致无融合生殖样表型。拟南芥开花调控基因FT(FLOWERING LOCUS T)是玉米中ZmZCN8(ZEA CENTRORADIALIS8)基因的同源基因,该基因编码ZmZCN家族的磷脂酰乙醇胺结合蛋白。在转基因植物中,ZmZCN8表达可导致早花,人为使用microRNA抑制ZmZCN8会导致晚花。INDETERMINATE1(ID1)基因编码单叶特异性锌指转录调控因子,该调控因子活化成熟玉米叶片中的ZmZCN8表达。通过对未成熟叶片的表观遗传调控,ID1蛋白调控ZmZCN8转录并促进核小体重塑。与ZMZCN8非常相似的ZmZCN7参与染色质的重塑调控,表明它可能编码不同的玉米成花素。氮(N)和磷(P)等土壤养分的摄入也受调控,生长在含氮较少环境中的植物可活化包括差异产生的miRNA的适应过程。miR169、miR399、miR408和miR528响应严重的低氮胁迫,而miRC10和miRC68可能响应土壤低氮胁迫。在硝酸盐缺失情况下,miR528s、miR169s、miR166s和miR408/b导致根系发育变化。miR169s、miR160f-5p、miR156s和miR171s在镉胁迫下分别通过靶向NFY、ARF、SBP和GRAS转录因子发挥作用。miR156负调控SBP盒转录因子tasselsheath4(tsh4),允许形成侧分生组织并抑制叶片起始,因此在建立分生组织和叶片边界中发挥着重要作用。miR164负调控NAC1表达,从而影响侧根生长。在玉米幼苗发育过程中,miRNA表达可以显示出显著差异。参与这一机制的大多数miRNA靶基因都参与转录调控。miR156、miR396、miR293和miR393分别靶向SPL、GRF、TIR1和LAC,并可能通过调控生长发育在调节籽粒灌浆中发挥关键作用。另外,miR164在种子发育过程中起调控作用,玉米籽粒的发育受9个保守的zma-miR169家族调控。
番茄
番茄(Solanum lycopersicum L.)是全球重要作物,是消耗量最大的蔬菜之一。由于甲基化抑制剂5-AzaC处理后表型发生逆转导致DNA甲基化,RNAi沉默MutS HOMOLOG1(MSH1)基因会影响生长和发育。嫁接介导的表观遗传变化用砧木中的MSH1系统检测,MSH1砧木突变阻碍siRNA形成。子代生长活力增强,可传递超过五代并依赖于RdDM。CG和CHG区域无色不成熟(cnr)基因启动子区域高甲基化使cnr沉默,导致成熟缺陷,在成熟期的野生果实中类似区域被去甲基化。2013年一项在番茄全基因组DNA甲基化研究中发现200多个成熟相关基因的启动子中5mC丢失,DNA甲基化下调与包括参与光合作用和细胞壁组织基因在内的成熟基因抑制有关。DEMETER样DNA去甲基化酶(DML)SlDML2在调控成熟番茄的DNA甲基化中起着关键作用,由于负责抑制负责果实成熟和软化基因的启动子区域甲基化增加,RNAi抑制SlDML2导致成熟缺陷。乙烯或类胡萝卜素通路中的许多差异甲基化基因编码转录因子和重要酶,这些转录因子和酶可能被不同产生的非编码RNA靶向。MSTRG.59396.1和miR396b靶向ACO2,MSTRG.43594.1和miR171b靶向CTR1; MSTRG.183681.1靶向ERF2,miR9470-3p靶向ERF5,MSTRG.95226.7靶向PSY1, 12:66127788|66128276靶向ZISO,MSTRG.181568.2靶向NCED,果实软化受组蛋白翻译后变化和染色质结构重塑的影响。
尽管不同亚家族基因可能发挥不同的作用,为调节番茄成熟,目前研究已经发现了多种组蛋白去乙酰化酶基因。RPD3/HDA1亚家族的SlHDA3和SlHDA1抑制参与细胞壁代谢基因,从而作为果实软化的负调控因子。另一方面,HD2亚家族的SlHDT3通过活化一组由SlHDA1/3调控的相关基因,在果实软化调控中发挥积极作用。SlERF.F12、TOPLESS 2(TPL2)和组蛋白去乙酰化酶HDA1/HDA3,下调了与成熟相关基因启动子区域的组蛋白乙酰化标记H3K9Ac和H3K27Ac,导致转录抑制。启动子中SINE反转录转座子的整合和插入的SINE高甲基化与维生素E(VTE3)基因表达下调有关。VTE3表达上调和果实VTE含量增加由VTE3启动子自发去甲基化导致。miRNA156通过靶向六个SQUAMOSA启动子结合蛋白(SBP)-box转录因子基因来调控植物高度、叶片大小和数量以及果实大小,miRNA156还通过靶向SPL转录因子调控番茄的卵巢和果实发育。miR157通过靶向LeSPL CNR基因调节番茄成熟,导致mRNA降解和翻译抑制。miR159以SGN-U567133基因为靶点,在叶和花的发育中发挥作用。miR167通过靶向ARF6和ARF8,导致花瓣、雄蕊和花柱较短,叶片大小和节间长度减小。miR4376通过靶向Ca2+-ATPase在番茄的生殖生长中发挥作用。miR168通过靶向AGO1而导致相变(phase transition)、叶片偏上性(leaf epinasty)和果实发育。
大豆
大豆(Glycine max (L.) Merr.)是一种重要的种子作物,可以用于饲料和人食用的蛋白质和油,目前共在45种不同的大豆品种中研究了表观遗传学在大豆驯化中的作用。DNA甲基化分析表明约75%的DMR由遗传变异以外的因素引起。2018年Raju等人报告了提高大豆产量和稳定性的育种系统,通过将野生型和MSH1获得的大豆记忆系杂交,利用MSH1系统在温室和田间试验中建立多种产量相关参数的epi-lines,所获得的表型几乎没有表型-环境互作,表明产量稳定性提高,受环境影响较小。MSH1抑制诱导的表观遗传变异至少可以传递三代,且可以通过多轮选择进行育种以提高和稳定作物产量。DNA甲基化是进化上保守的修饰,特别是在对称CG环境中,会导致影响叶片形态、开花期、花器官特性、生育力和胚胎发生的基因沉默。同时种子发育还受ATP依赖性SWI2/SNF2染色质重塑因子met1和ddm1突变的影响,大豆和DMR中显著的CG甲基化发生在子叶、叶、茎和根的甲基化组之间。2017年An等人报告了DNA甲基化在种子发育过程中的作用,在子叶CG(66%)、CHG(45%)和CHH(9%)区域观察到甲基化,CHH甲基化水平从初始阶段的6%急剧增加到后期阶段的11%,而转录基因的甲基化差异是非转录基因的两倍;在CG、CHG和CHH环境中,共鉴定出40、66和2136个DMR基因,且它们的表达与甲基化之间呈负相关。CHH环境下种子成熟期间的大多数DMR基因转录下调,且与DNA复制和细胞分裂相关。在大豆种皮中,DNA甲基化导致调节花青素合酶基因的MYB转录因子的TE元件的转座和剪接发生变化。
油菜
油菜(Brassica napus L.)也是一种主要的油籽作物,是富含蛋白质的牲畜饲料的主要来源。能量消耗效率是植物产量和活力的显著特征,具有可受人工选择影响的表观遗传成分。等基因油菜群体中产生的具有不同生理和农艺性状群体,可以重复选择单植株及其自交子代作为呼吸强度。如重复选择甘蓝型油菜等基因系可产生稳定的表观基因型,具有较高的能量利用效率。将这些表观遗传不同但遗传序列相同的品系进行杂交,促进杂交品种产量提高了5%。BrSDG8突变(编码组蛋白甲基转移酶,影响开花位点C染色质中的H3K4三甲基化)与早期抽薹相关,其调控着叶头形成和种子产量等性状。在另一项研究中,Bra032169基因(编码组蛋白甲基转移酶)的插入突变与早期抽薹有关。最近报道了Jumonji H3K27me3去甲基化酶(BraA.REF6和BraA.ELF6)在调控花转变中的作用,在调控H3K27me3水平方面,BraA.REF6比BraA.ELF6更重要。在braA.ref6突变中,FLC基因表达保持不变,开花延迟。而在braA.elf6突变的FLC中,H3K27me3水平较高,导致开花提前。通过上游FLC组分的表观遗传变化,BrcuHAC1可能影响抽薹和开花时间。去甲基化药物5-AzaC导致目标区域的DNA低甲基化,从而促进种子蛋白和亚油酸含量增加。2012年Solis等人发现DNA甲基化下调可以启动小孢子胚胎发生,细胞特异性DNA甲基化增加与花粉和胚胎分化有关。5-AzaC可以增加小孢子重编程、全能获得和胚胎发生起始,并阻止胚胎分化,表明甲基化在抑制小孢子重编程和全能获得中的作用。新型小分子BIX-01294可降低组蛋白H3K9甲基化,从而促进小孢子重编程和胚胎发生的启动。在细胞重编程和胚胎发育过程中,H3K9me2和HKMT参与胚胎细胞分化和异染色质化,而H3Ac、H4Ac和HAT参与转录活化、全能和增殖。
其他与农艺性状相关的作物表观遗传变化
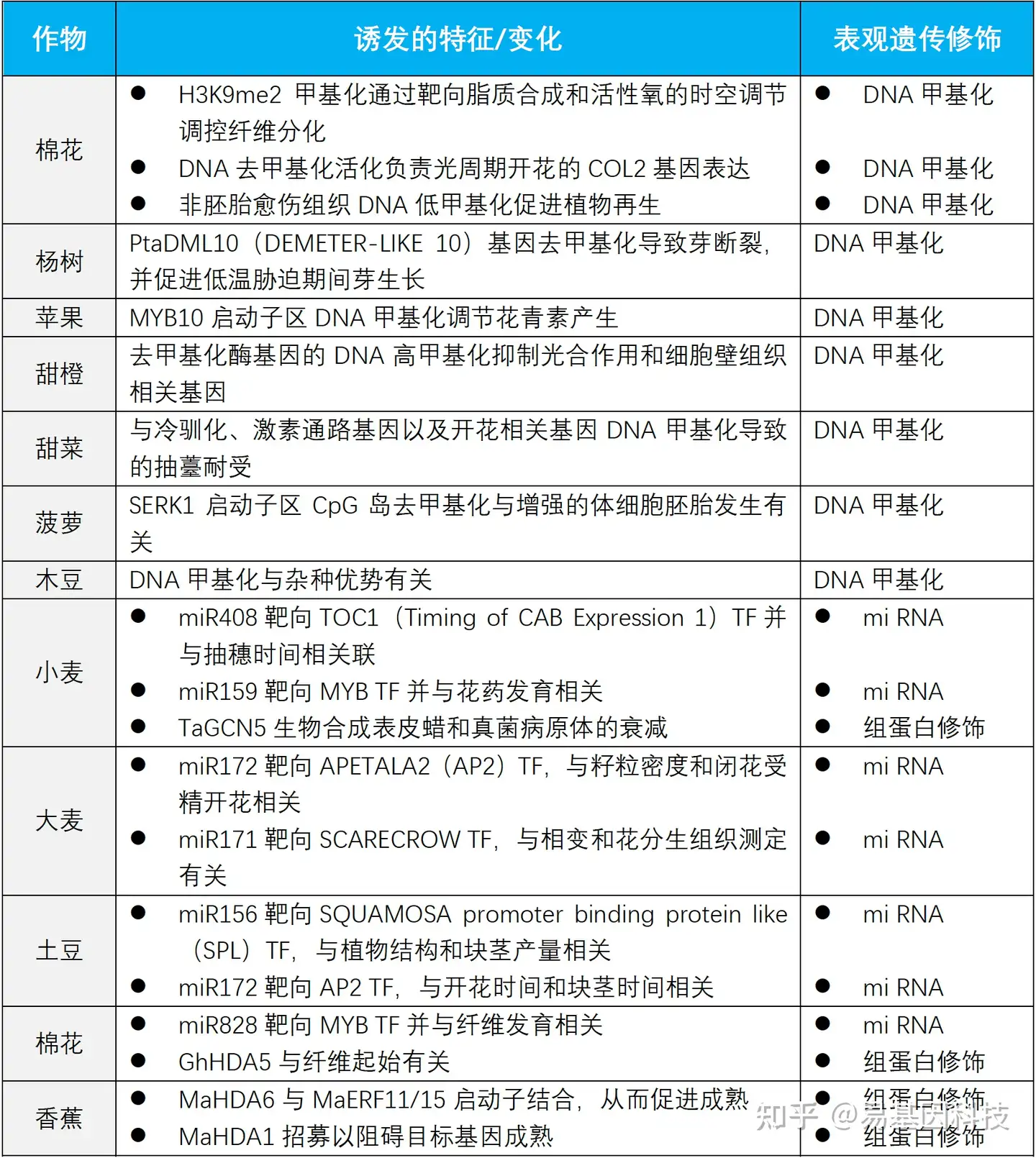
作物表观遗传学和表观育种前景展望
表观遗传学促进表型变异,因此了解表观遗传学和表观基因组学将有助于阐明环境因素影响植物表型机制。育种家可能通过使用现有的表观遗传变化或表观基因组修饰来解决遗传侵蚀的持续问题并揭示隐性变异。Epibreeding育种计划可以在一代中产生广泛的表型变异,一些变化可能从一代传递到下一代,最终克服所有降低作物育种计划有效性的所有制约因素。新的表观等位基因(epialleles)在遗传多样性较低的种群尤为重要。由于目前育种技术主要集中在遗传学上而忽略了表观遗传因素,因此在epialleles水平上利用表观遗传信息可能为作物发育提供新的机会。当然,需要对更多更广泛的植物物种进行进一步研究,以更深入地了解触发和维持作物表观遗传多样性的过程。
易基因科技提供全面的表观组学测序服务,有DNA甲基化、组蛋白修饰、RNA甲基化测序需要的老师可联系。
参考文献:Gupta C, Salgotra RK. Epigenetics and its role in effecting agronomical traits. Front Plant Sci. 2022;13:925688.
更多阅读:
作物育种:MdMTA介导的RNA甲基化(m6A修饰)在苹果抗逆品种选育中的作用研究
3文聚焦:植物逆境胁迫响应的表观遗传调控机制(抗干旱+耐高温+抗干耐盐)
3文聚焦:DNA甲基化在动物育种中的作用研究成果(家蚕+绵羊+肉鸡)
综述:表观遗传学和表观育种在作物品种改良的重要作用(水稻+玉米+番茄+大豆+油菜)
标签:DNA,综述,调控,组蛋白,基因,甲基化,品种改良,表观 From: https://www.cnblogs.com/E-GENE/p/16788686.html