大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
研究人员已经在多种植物物种中观察到在杂交过程中跨染色体间互作而导致DNA甲基化变化。然而,这些互作的原因或结果知之甚少。
2023年12月18日,佛罗里达大学微生物学和细胞科学系meixia Zhao等人在《Plant Physiology》杂志发表题为“Heritable changes of epialleles near genes in maize can be triggered in the absence of CHH methylation”的研究论文,通过WGBS和对应的RNA-seq等分析揭示了杂交植物玉米(Zea mays)中DNA甲基化模式的整体变化,且这些变化与小RNA数量变化无关。

标题:Heritable changes of epialleles near genes in maize can be triggered in the absence of CHH methylation(CHH甲基化丢失可触发玉米表观等位基因的可遗传变化)
时间:2023-12-18
期刊:Plant Physiology
影响因子:7.4 / 1区
技术平台:WGBS、RNA-seq、sRNA-seq等
研究摘要:
本研究比较了玉米(Zea mays)中小RNA生物发生基因Mop1(paramutation介导因子1)突变体的杂交种一代(F1代)的DNA甲基化组,以及与其亲本、野生型同胞和玉米回交后代的DNA甲基化组。研究数据表明,杂交介导了跨染色体甲基化(trans-chromosomal methylation,TCM)和跨染色体去甲基化(trans-chromosomal demethylation,TCdM)的整体变化,其中大部分是CHH甲基化变化。在这些可获得小RNA的TCM差异甲基化区域(DMR)中,超过60%的区域没有观察到小RNA数量的显著变化。在mop1突变体中,CHH TCM DMRs的甲基化大部分丢失,尽管这种突变体的作用依赖于其DMRs位点。且TCM DMR中CHH的增加与部分高表达基因表达增强和少量低表达基因表达抑制相关。回交植物中甲基化水平检测结果表明,TCM和TCdM都可以在下一代中维持,但TCdM比TCM更稳定。令人惊讶的是,尽管F1植物中大多数TCM DMR中CHH甲基化增加需要Mop1,但这些DMR新表观遗传状态启动不需要该基因的功能拷贝,表明这些变化的启动与RNA介导的DNA甲基化无关。
简而言之,这项研究揭示了杂交植物中DNA甲基化模式的整体变化,并且这些变化与小RNA的数量变化无关。此外,Mop1基因在维持某些甲基化模式中起着重要作用,但新表观遗传状态的建立并不依赖于这个基因,意味着DNA甲基化变化可能还有其他未知的调控机制。
材料方法:
在Mo17背景下的mop1杂合子玉米(Z. mays)植物与B73背景下的mop1杂合子植物(Mo17;mop1-1/+× B73;mop1-1/+)杂交以产生F1杂交mop1突变体(Mo17/B73;mop1/mop1)及其杂交野生型同胞(Mo17/B73;+/+;图1A)。Mop1突变至少已经在B73和Mo17下传递了7代。收集F1植株和两个亲本系(B73和Mo17)植株5~7cm的未成熟玉米穗用于后续的全基因组重亚硫酸盐测序(WGBS)、RNA测序(RNA-seq)和小RNA测序(sRNA-seq),所有样本均设置两个生物学重复。
研究结果:
(1)杂交植物中CHH甲基化水平在全基因组范围内增加
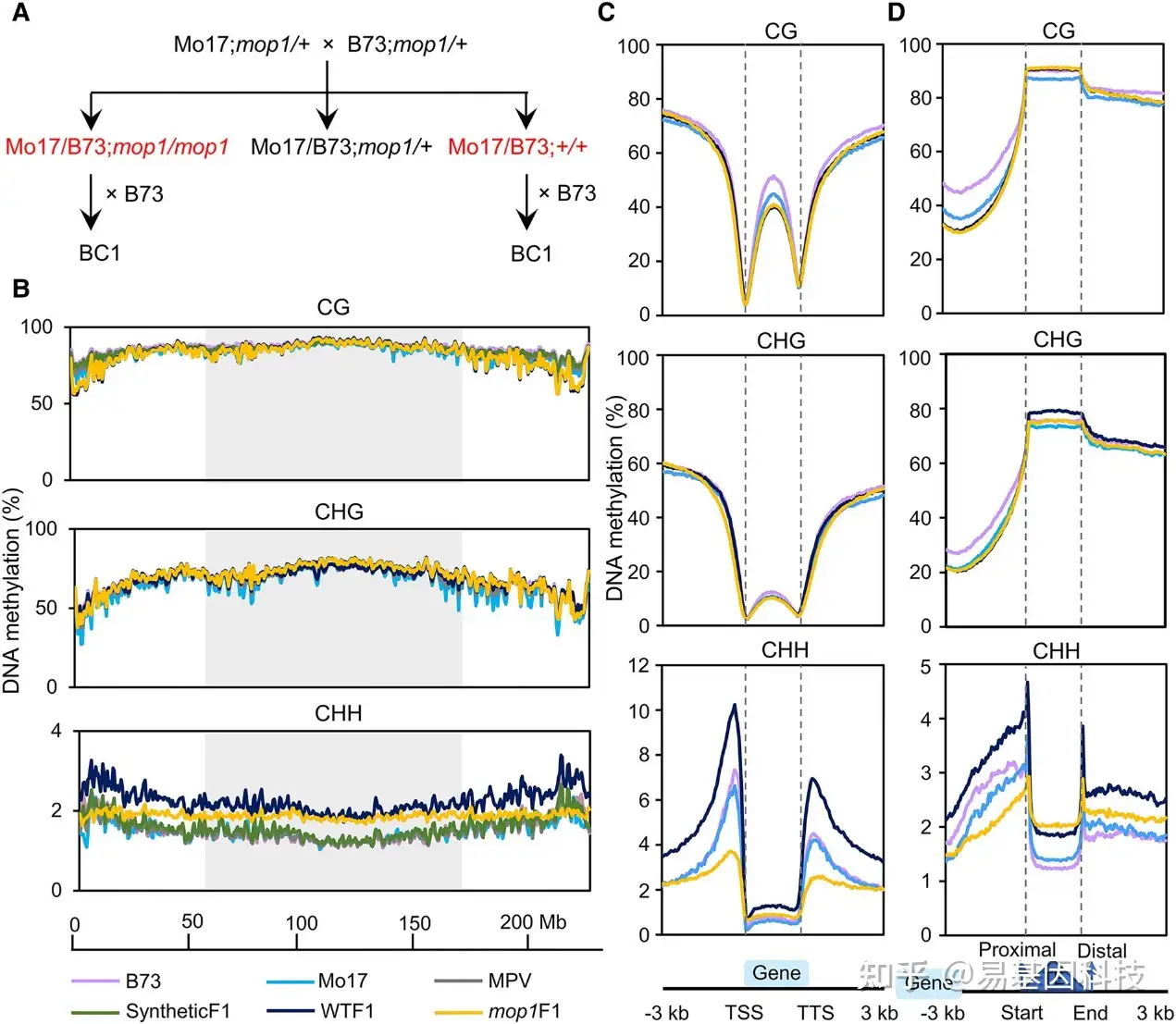
图1:杂交植物中CHH甲基化水平在全基因组范围内增加。
- 构建野生型F1(WTF1)、mop1突变体F1(mop1F1)和BC1的遗传策略。
- CG、CHG和CHH甲基化在5号染色体上的分布。甲基化水平在1 Mb windows中检测,每个窗口以500 kb移位。阴影框代表着丝粒周围区域。MPV表示midparent值。合成的F1杂交甲基化组是通过B73和Mo17亲本相等数量的WGBS reads作为对照而混合生成。
- 基因(gene)中和侧翼区域的甲基化模式。
- 转座元件(TE)中和侧翼区域的甲基化模式。
DNA甲基化水平在基因/TEs ±3 kb区域内的50 bp窗口中计算。每个基因/TE序列被分成40个等大小的Bin来检测基因/TE body甲基化。由于基因/TEs长度不同,Bin大小在不同的基因/TEs之间有所不同。根据TEs侧翼基因的不同,TEs甲基化被定向为近端和远端。每个样本的甲基化水平是计算为每个序列上下文中甲基化C的比例除以总C的比例(CG、CHG和CHH,其中H = A、T或C),并对每个窗口进行平均。通过结合每个基因型的2个生物学重复来确定平均甲基化水平。
(2)在F1杂交的TCM DMR中,高亲本和低亲本(甲基化水平较高和较低的亲本)等位基因的CHH甲基化水平均升高
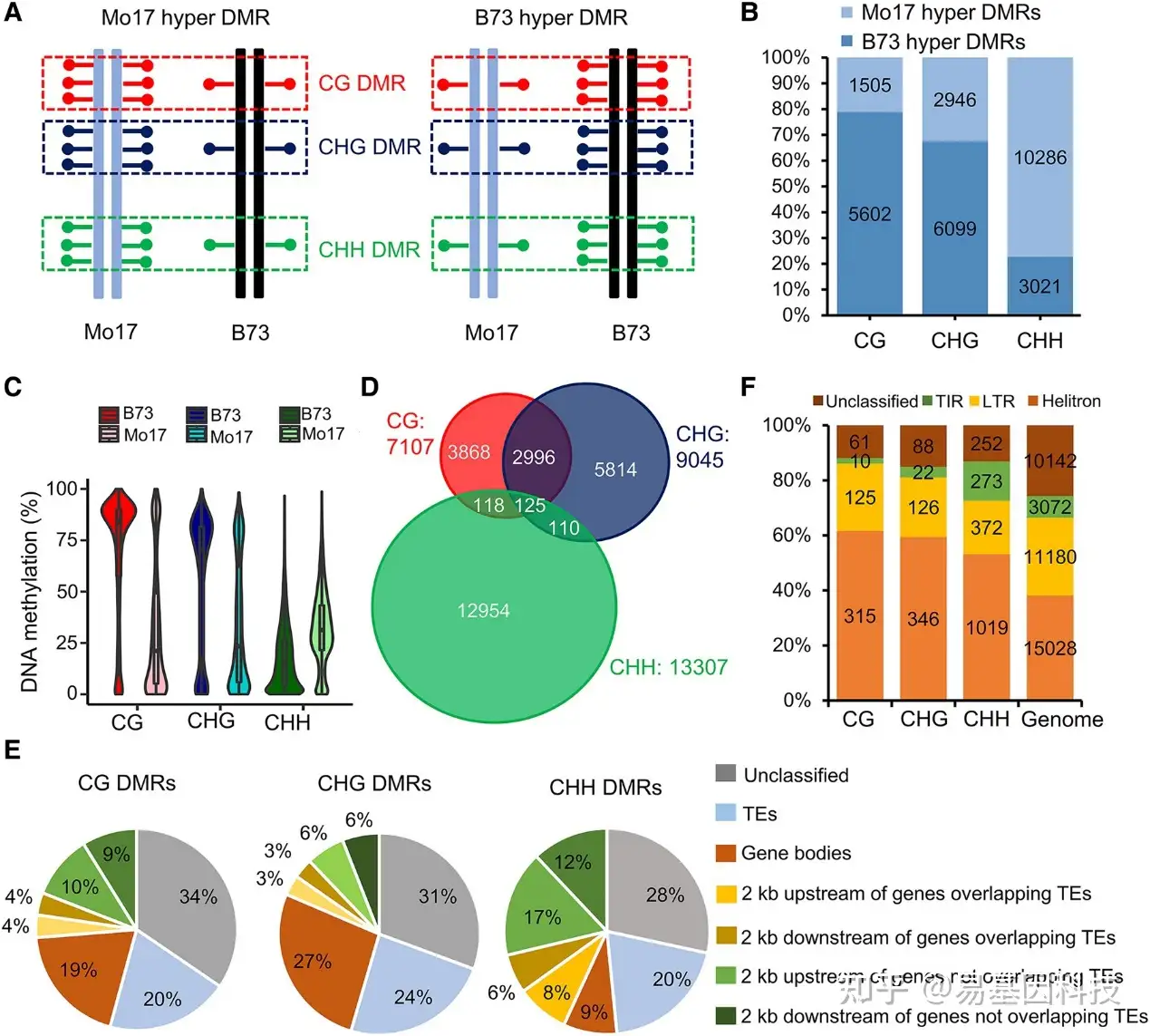
图2:亲本CHH DMR主要位于基因的±2kb侧翼区域。
- 亲本B73高DMR(B73中较高甲基化)和Mo17高DMR(Mo17中较高甲基化)的定义。红色(上)、蓝色(中)和绿色(下)点分别代表CG、CHG和CHH甲基化。
- B73具有更多的CG和CHG高DMR,而Mo17具有更多的高CHH DMR。
- B73在CG和CHG DMR处具有较高的甲基化水平,而Mo17在CHH DMR处具有较高的甲基化水平。箱形图底部和顶部边界是第一和第三四分位数,箱体内粗线是中位数,称为第二四分位数。线条两端代表数据的最小值和最大值。
- CG和CHG DMR之间的重叠比其与CHH DMR之间的重叠更多。
- CG、CHG和CHH亲本DMR分布;在基因上下游 ±2kb区域内与TEs重叠DMR表明DMR与基因侧翼区域内的TE重叠。
- 在E中,基因上下游 ±2kb区域内与TE重叠的TE DMR类型。DMR,差异甲基化区域。
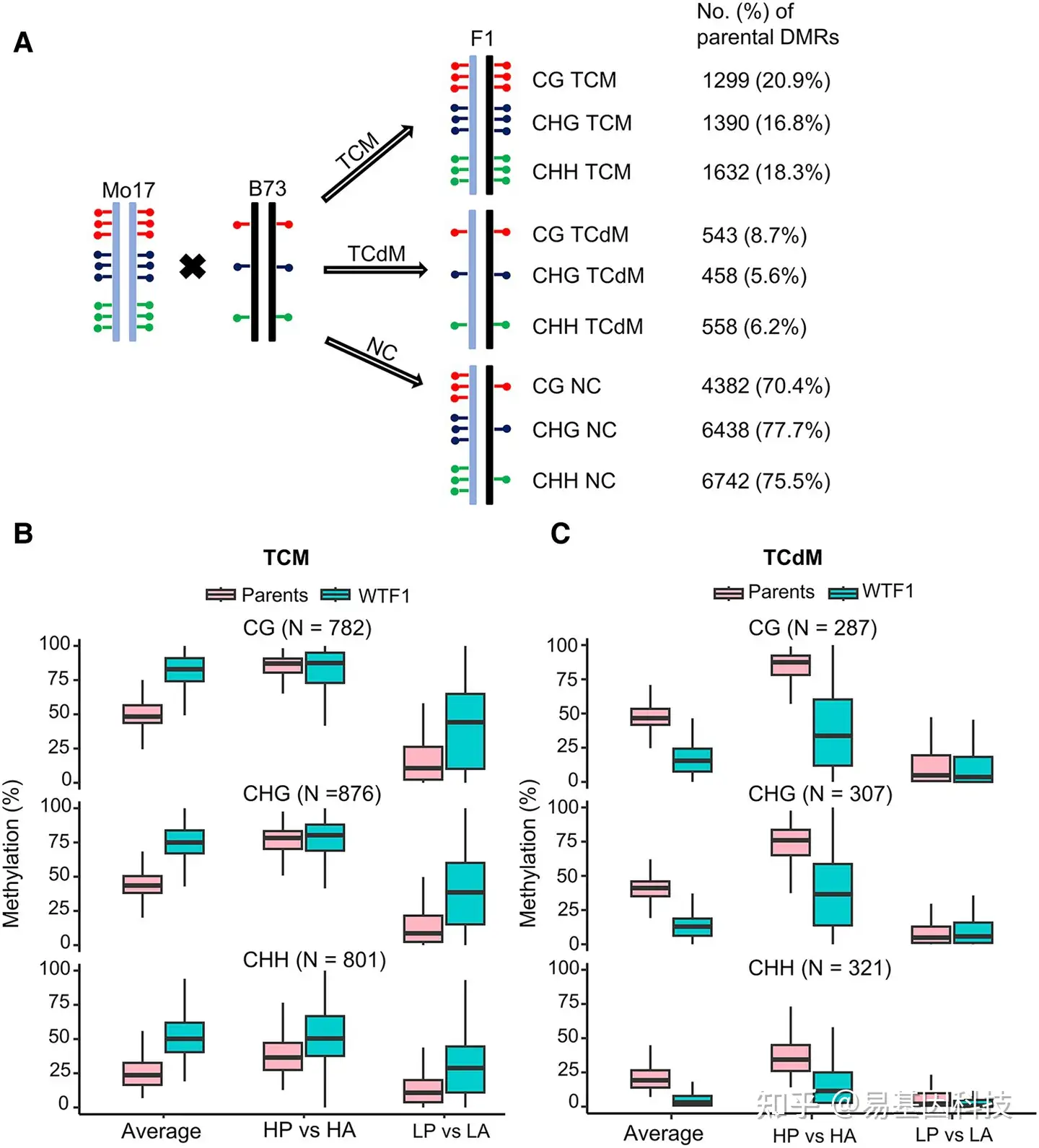
图3:在F1杂交的TCM DMR中,高亲本和低亲本等位基因的CHH甲基化水平均增加。
- 鉴定WTF1和亲本之间的TCM、TCdM和未改变(NC)DMR。
- 亲本和WTF1中TCM DMR的CG、CHG和CHH甲基化的比较。
- 亲本和WTF1中TCdM DMR的CG、CHG和CHH甲基化的比较分析。
对于B)和C),箱形图的底部和顶部边界是第一和第三四分位数,单个box内的粗线是中位数,称为第二四分位数。Box线两端代表数据的最小值和最大值。HP,高亲本(甲基化水平较高的亲本);HA,F1中的高亲本等位基因;LP,低亲本(甲基化水平较低的亲本);LA,F1中的低亲本等位基因。平均值是指WTF1和mop1F1中两个亲本之间或两个等位基因之间的平均值。DMR,差异甲基化区域;TCM,跨染色体甲基化;TCdM,跨染色体去甲基化。
(3)mop1突变体中CHH TCM DMR的甲基化显著降低
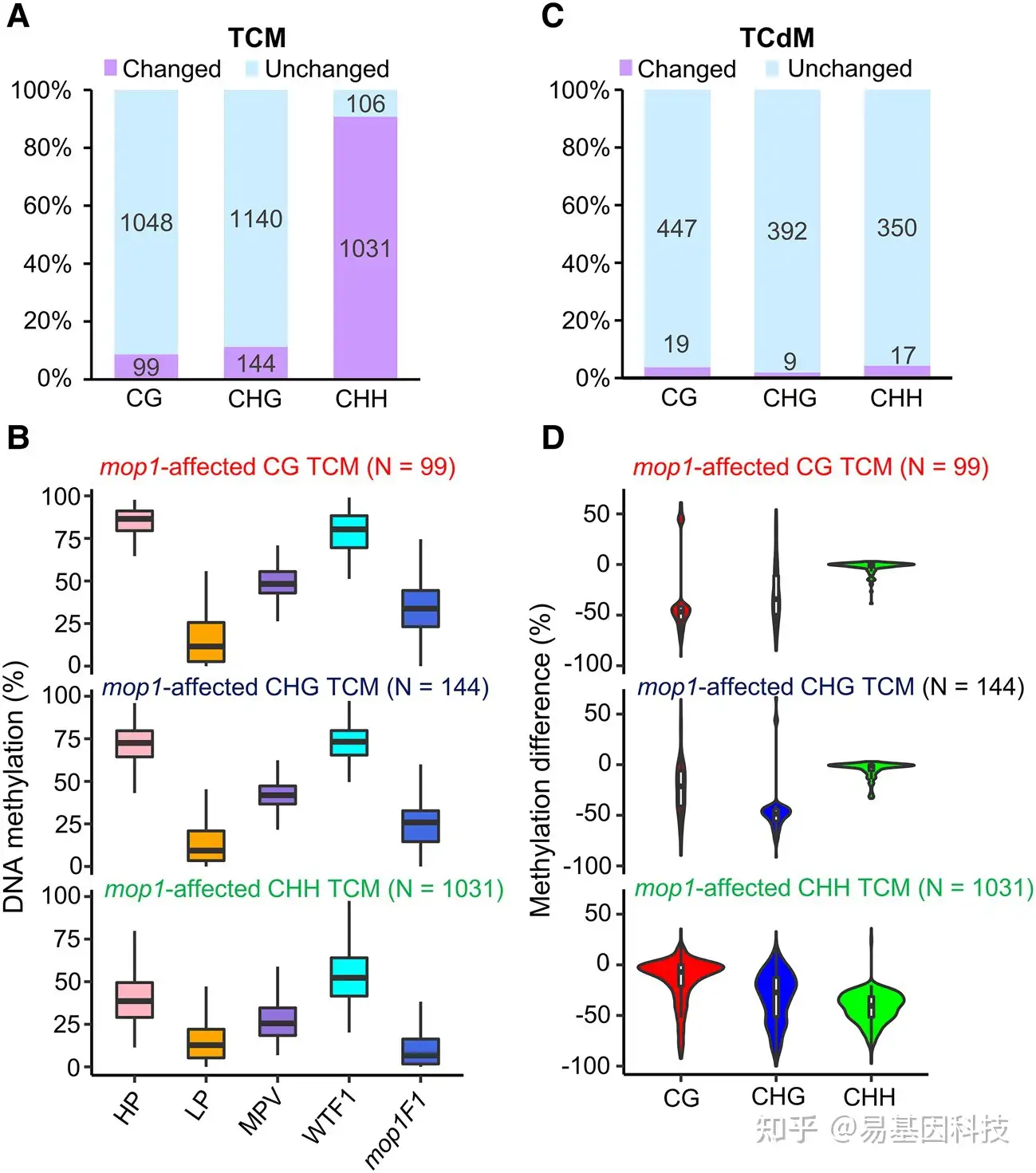
图4:mop1突变主要去除了CHH TCM DMR的甲基化。
- mop1突变影响的CG、CHG和CHH TCM DMR的数量。
- 比较mop1影响CG、CHG和CHH TCM DMR的甲基化水平。
- mop1突变影响的CG、CHG和CHH TCdM DMR的数量。
- 在mop1影响的CG、CHG和CHH TCM DMR中检测其他2种胞嘧啶背景下的甲基化变化。上图显示了99个受mop1影响的CG TCM DMR的CHG和CHH序列背景下的甲基化变化。中间面板显示了144个受mop1影响的CHG TCM DMR的CG和CHH序列背景下的甲基化变化。下图显示了1031个受mop1影响的CHH TCM DMR的CG和CHG序列背景下的甲基化变化。
(4)1个等位基因小RNA足以触发F1杂交植物中大多数CHH TCM DMR中其他等位基因甲基化
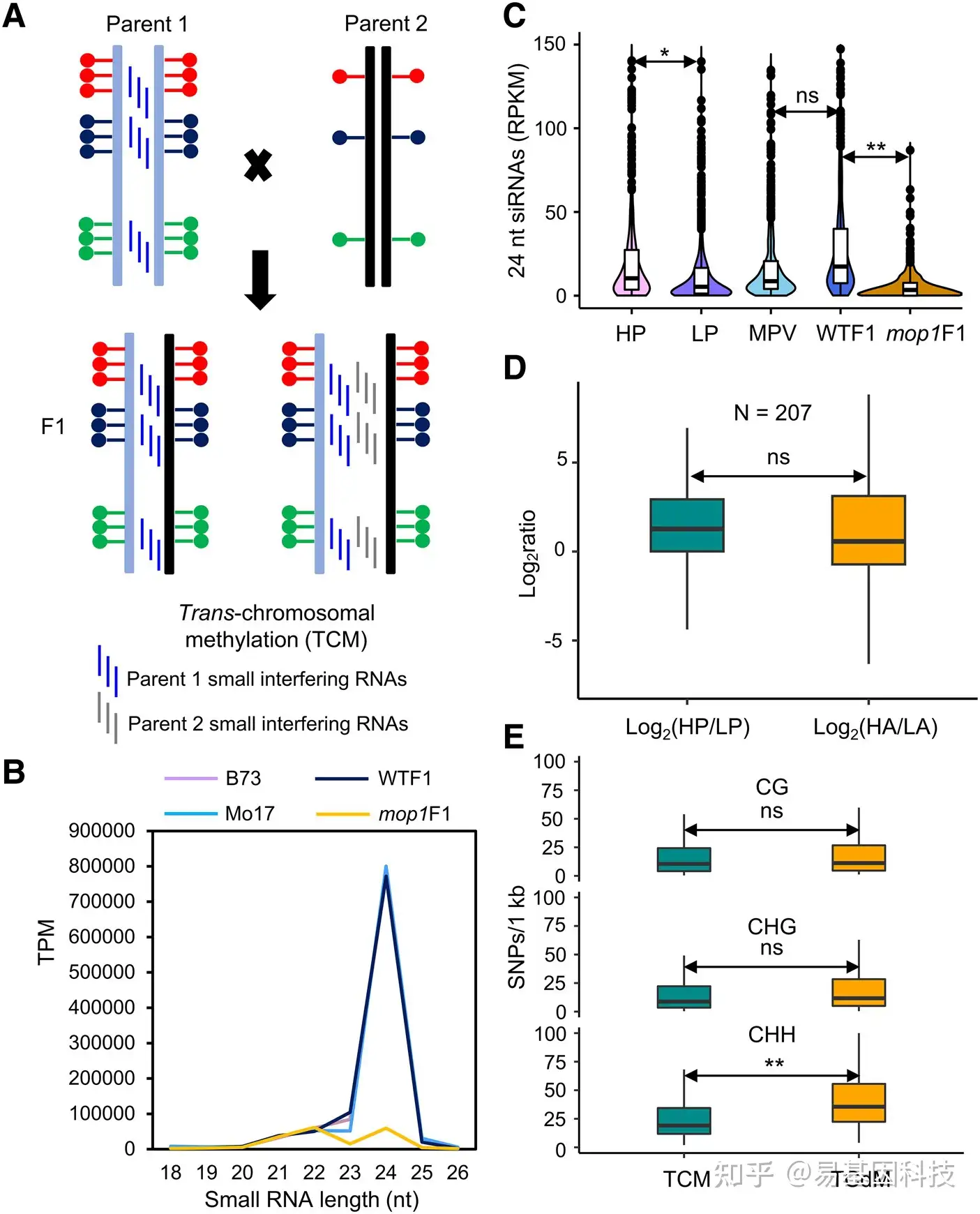
图5:从一个亲本产生的小RNA足以触发杂交后代中其他等位基因的新甲基化。
- F1代CHH TCM中小RNA生物发生的两个假设模型。
- 亲本、WTF1和mop1F1中小RNA的表达值。TPM,唯一比对reads/M的转录本。根据先前研究,将小RNA值调整为所有成熟microRNA的总丰度。
- mop1上24-nt siRNA的丰度影响CHH TCM DMR。HP,高亲本(甲基化水平较高的亲本);LP,低亲本(甲基化水平较低的亲本);MPV,中亲值。
- 在受mop1影响的CHH TCM DMR中,高亲本与低亲本以及高亲本等位基因与低亲本等位基因的24 nt siRNA的比率。
- TCM和TCM之间的SNP数量。
晶须之外的单个数据点是潜在的异常值,表明值与数据的整体模式显著偏离。**P<0.01和*P<0.05,学生t检验。ns,不显著;DMR,差异甲基化区域;TCM,跨染色体甲基化;TCdM,跨染色体去甲基化。
(5)侧翼基因序列的CHH甲基化可能与邻近基因表达抑制或增强有关
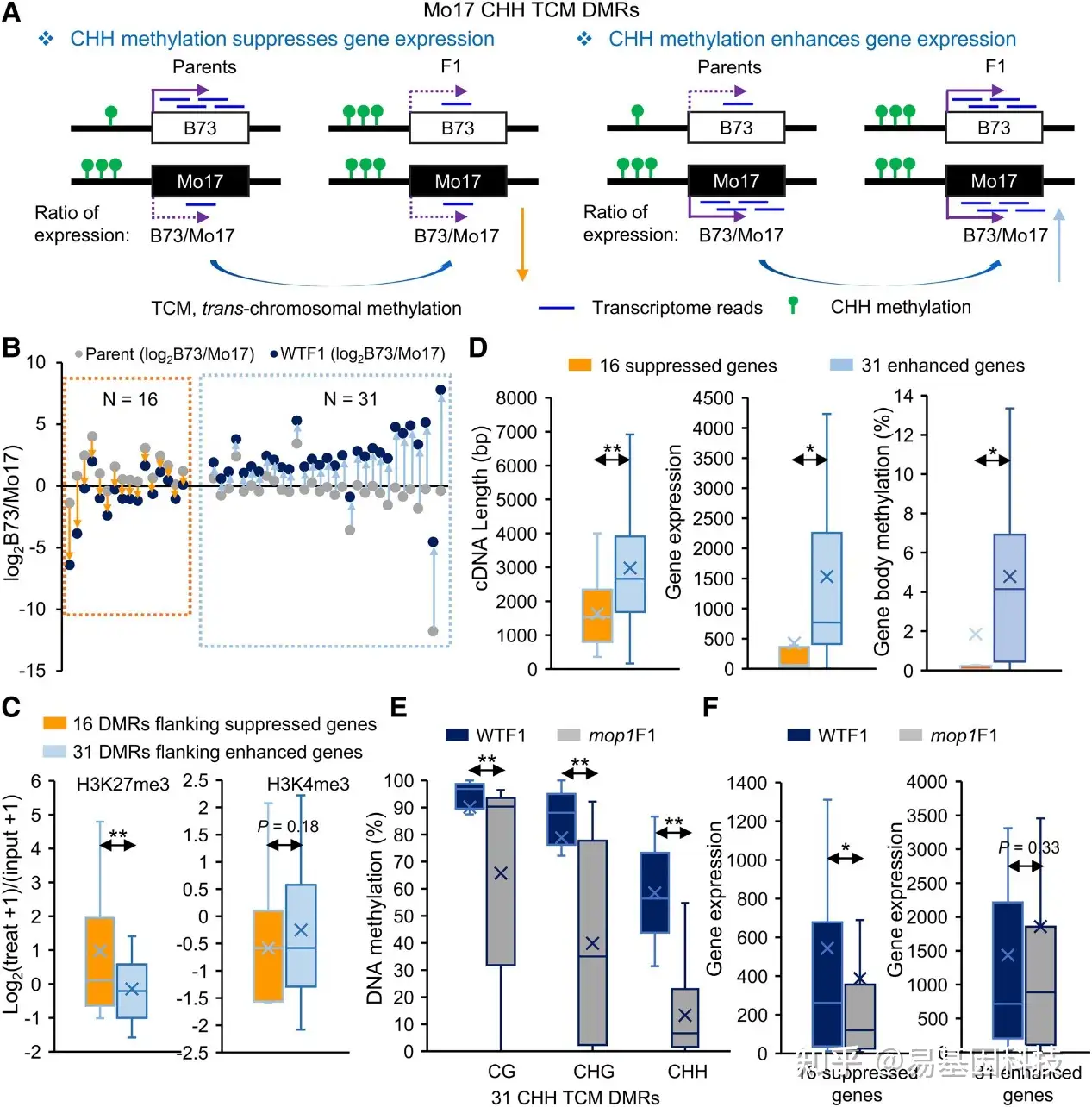
图6:CHH甲基化与邻近基因的抑制和增强表达有关。
- CHH甲基化对基因表达影响的两种可能情况。仅显示了Mo17 CHH TCM DMR的例子。紫色(水平)实心和虚线箭头表示该基因在亲本或等位基因中的表达增强或抑制。橙色(向下)和蓝色(向上)箭头表示B73与Mo17的比率降低和增加。
- 根据A)中的模型鉴定出的基因数量。亲本和WTF1之间B73与Mo17表达值的比率用于区分。
- 与邻近基因表达抑制相关的16个CHH-TCM DMR和与邻近基因表达增强相关的31个CHH-TCM DMR的组蛋白修饰。
- 16个抑制基因和31个增强基因的基因特性。去除内含子以检测基因体甲基化。
- 与邻近基因表达增强有关的31个CHH TCM DMR处mop1突变体的甲基化变化。**P<0.01,学生配对t检验。
- mop1突变体中16个抑制基因和31个增强基因的基因表达变化。
(6)启动CHH TCM位点的表观遗传状态变化不依赖于MOP1
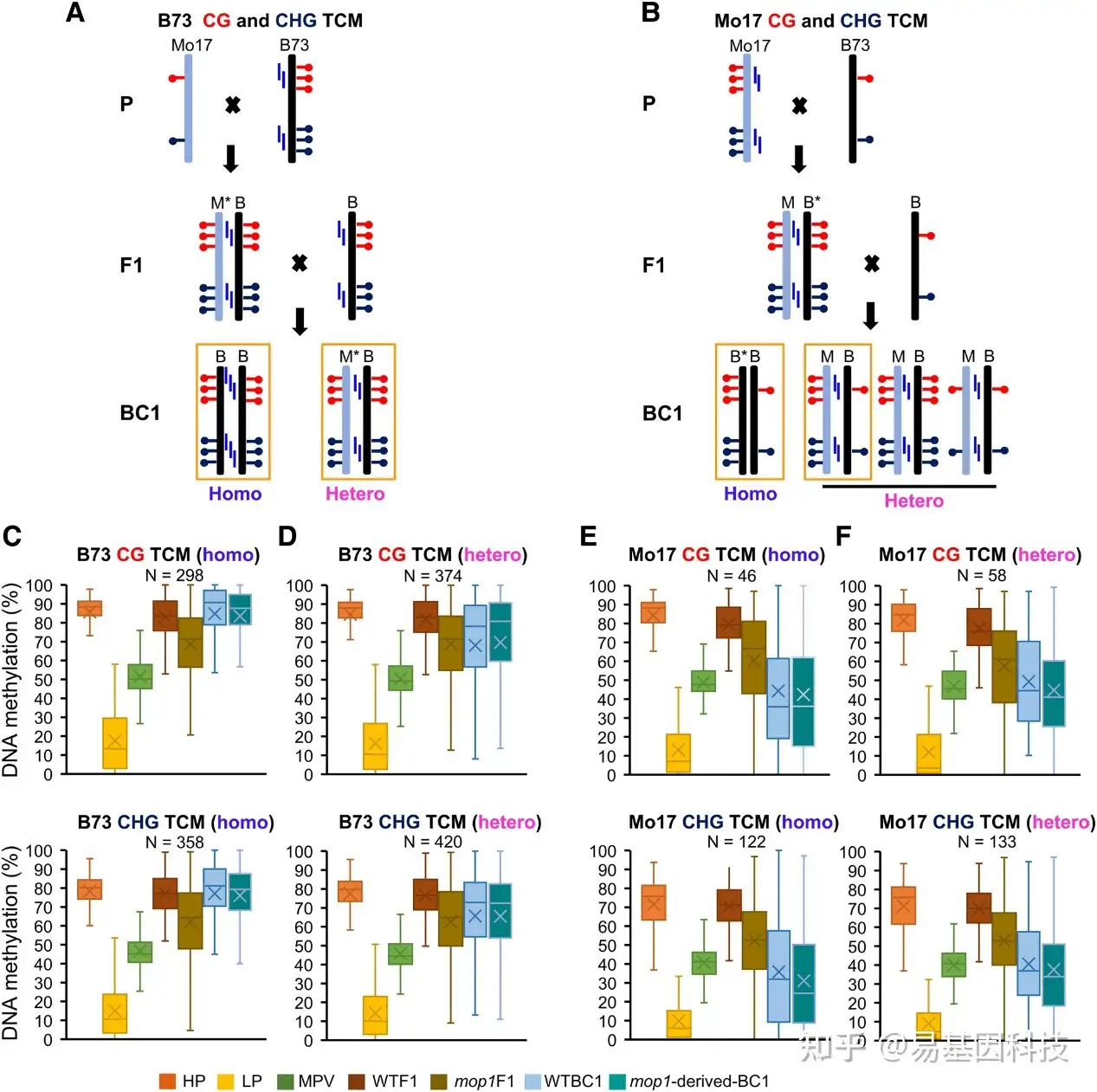
图7:F1植物中CG和CHG 跨染色体甲基化(TCM)DMR的新触发甲基化在下一代得以维持。
- B73 CG和CHG TCM DMR维持的假设模型。星号(M*)表示新转换的(甲基化)等位基因。M、 Mo17;B、 B73。每个等位基因附近的蓝色垂直条表示siRNA,橙色框表示C到F中数据支持的模型。
- Mo17 CG和CHG TCM DMR维持的假设模型。星号(B*)表示新转换的(甲基化)等位基因。M、 Mo17;B、 B73。
- 纯合子B73 CG和CHG TCM DMR的甲基化变化。
- 杂合子B73 CG和CHG TCM DMR的甲基化变化。
- 纯合子Mo17 CG和CHG TCM DMR的甲基化变化。
- 杂合子Mo17 CG和CHG TCM DMR的甲基化变化。
C-F箱形图的底部和顶部边界是第一和第三四分位数,单个箱形图的粗体线是中位数,称为第二四分位数。晶须的末端(线)代表数据的最小值和最大值。DMR,差异甲基化区域; TCM,跨染色体甲基化;Homozygous,纯合子;Heterozygous,杂合子;WTBC1,Mo17/B73;+/+ × B73;mop1衍生的BC1,Mo17/B73;mop1/mop1 × B73。
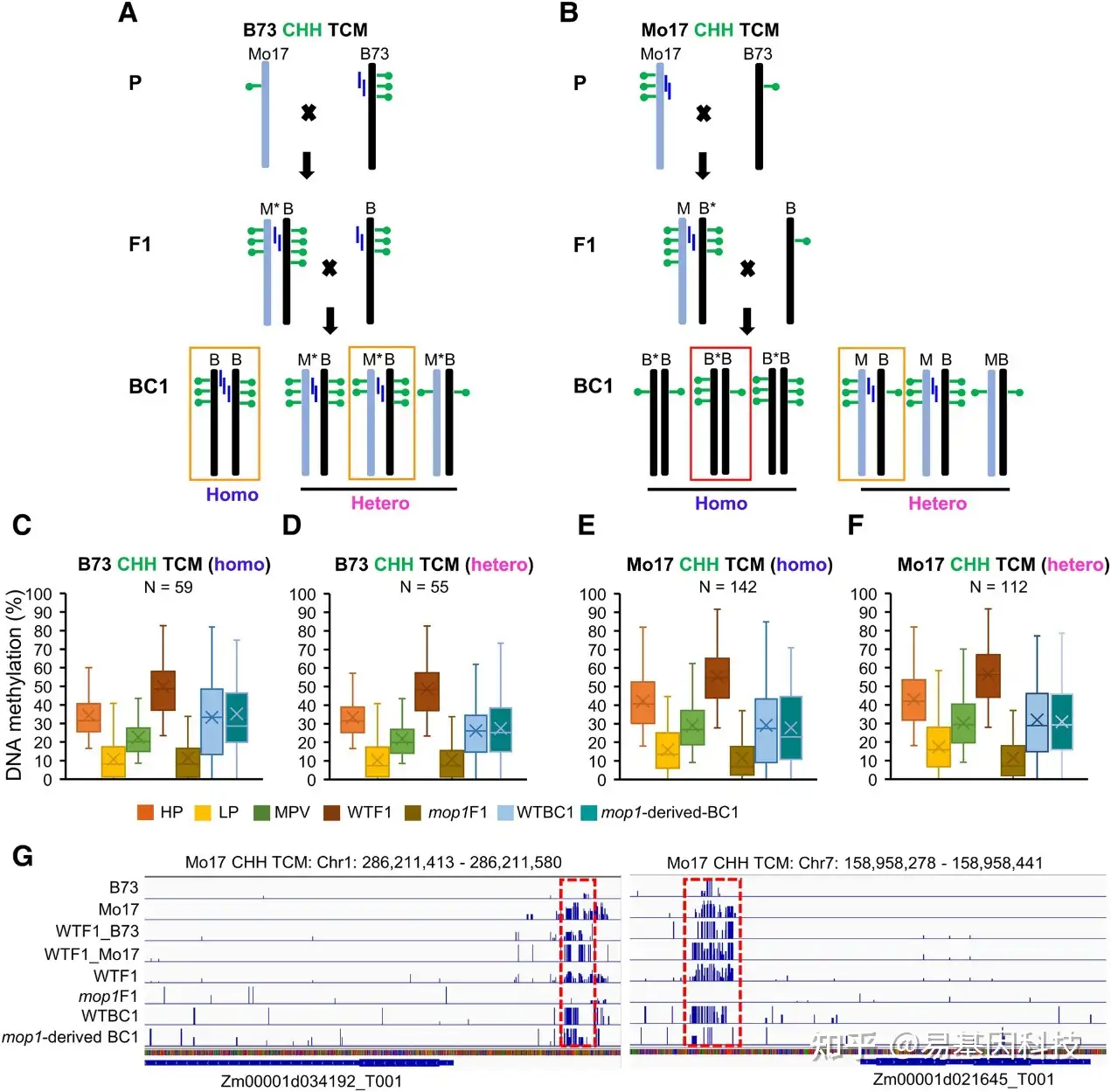
图8:玉米中跨染色体CHH甲基化的表观遗传状态变化启动不依赖于MOP1。
- B73 CHH TCM DMR维持的假设模型。星号(M*)表示新转换的(甲基化)等位基因。M、 Mo17;B、 B73。每个等位基因附近的蓝色垂直条代表siRNA。
- Mo17 CHH TCM DMR维持的假设模型。星号(B*)表示新转换的(甲基化)等位基因。M、 Mo17;B、 B73。橙色(从左到右,编号为1、2和4的box)和红色(编号为3)box表示C到F中数据支持的模型。
- 纯合子B73 CHH TCM DMR的甲基化变化。
- 杂合子B73 CHH TCM DMR的甲基化变化。
- 纯合子Mo17 CHH TCM DMR的甲基化变化。
- 杂合子Mo17 CHH TCM DMR的甲基化变化。
- 2个Mo17 CHH TCM DMR的分布和甲基化水平示例。红色虚线框突出显示每个示例中的DMR。
关于易基因全基因组重亚硫酸盐测序(WGBS)
全基因组重亚硫酸盐甲基化测序(WGBS)可以在全基因组范围内精确的检测所有单个胞嘧啶碱基(C碱基)的甲基化水平,是DNA甲基化研究的金标准。WGBS能为基因组DNA甲基化时空特异性修饰的研究提供重要技术支持,能广泛应用在个体发育、衰老和疾病等生命过程的机制研究中,也是各物种甲基化图谱研究的首选方法。
易基因全基因组甲基化测序技术通过T4-DNA连接酶,在超声波打断基因组DNA片段的两端连接接头序列,连接产物通过重亚硫酸盐处理将未甲基化修饰的胞嘧啶C转变为尿嘧啶U,进而通过接头序列介导的 PCR 技术将尿嘧啶U转变为胸腺嘧啶T。
应用方向:
WGBS广泛用于各种物种,要求全基因组扫描(不错过关键位点)
- 全基因组甲基化图谱课题
- 标志物筛选课题
- 小规模研究课题
技术优势:
- 应用范围广:适用于所有参考基因组已知物种的甲基化研究;
- 全基因组覆盖:最大限度地获取完整的全基因组甲基化信息,精确绘制甲基化图谱;
- 单碱基分辨率:可精确分析每一个C碱基的甲基化状态。

易基因科技提供全面的DNA甲基化研究整体解决方案,详询易基因:0755-28317900。
参考文献:Liu B, Yang D, Wang D, Liang C, Wang J, Lisch D, Zhao M. Heritable changes of epialleles near genes in maize can be triggered in the absence of CHH methylation. Plant Physiol. 2023 Dec 18. pii: 7477629. doi: 10.1093/plphys/kiad668. PubMed PMID: 38109503.
相关阅读:
年终盘点 | 易基因2023年度DNA甲基化研究项目文章精选
项目文章 | WGBS+RNA-seq揭示黄瓜作物的“源-库”关系受DNA甲基化调控
项目文章 | WGBS等揭示SOX30甲基化在非梗阻性无精症中的表观遗传调控机制
项目文章|WGBS+RNA-seq揭示PM2.5引起男性生殖障碍的DNA甲基化调控机制
原文阅读:
作物研究:CHH甲基化丢失可触发玉米表观等位基因的可遗传变化
标签:Mo17,CHH,等位基因,甲基化,DMR,TCM,亲本 From: https://www.cnblogs.com/E-GENE/p/18036724