大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
视网膜母细胞瘤(retinoblastoma, RB)是儿童期最常见的眼内恶性肿瘤,可导致失明甚至死亡。RB1缺失(>90%)和MYCN扩增(~10%)被认为是致癌驱动事件,导致细胞周期更新增强和癌基因激活。最近的研究表明,表观遗传缺陷也参与RB肿瘤进展。NSUN2介导的N5-甲基胞嘧啶(m5C)修饰通过激活癌基因或抑制肿瘤抑制因子参与多种肿瘤的细胞增殖和侵袭。m5C在RNA代谢中起重要作用,对于维持表观基因组稳态至关重要。然而,m5C修饰在致癌过程中的作用尚未完全明确。
2023年5月25日,上海交通大学医学院第九人民医院眼科陆琳娜、柴佩韦、范佳燕等共同通讯在《Clin Transl Med》期刊发表题为“NSUN2-mediated m5 C RNA methylation dictates retinoblastoma progression through promoting PFAS mRNA stability and expression”的研究论文,探讨了NSUN2介导的m5C RNA甲基化如何通过促进磷酸核糖甲酰甘氨酸脒合酶(phosphoribosylformylglycinamidine synthase,PFAS) mRNA的稳定性和表达来影响视网膜母细胞瘤(RB)的进展。

标题:NSUN2-mediated m5C RNA methylation dictates retinoblastoma progression through promoting PFAS mRNA stability and expression(NSUN2介导的m5C RNA甲基化通过促进PFAS mRNA稳定性和表达来决定视网膜母细胞瘤的进展)
时间:2023.5.25
期刊:Clin Transl Med
影响因子:IF 10.6
实验方法:m5C甲基化RNA免疫沉淀测序(m5CmeRIP-seq)等
研究摘要:
本研究通过视网膜母细胞瘤(RB)细胞和临床样品中的mRNA分离和抗m5C dot blot试验检测全局和mRNA m5C水平。建立原位眼内异种移植模型以检查RB的致癌行为,病进行全基因组多组学分析以鉴定NSUN2的功能靶标,包括蛋白质组学分析,转录组筛选和m5C甲基化RNA免疫沉淀测序(m5C meRIP-seq)。此外进行了基于类器官的单细胞分析和基因相关性分析,以验证NSUN2/ALYREF/m5C PFAS致癌级联反应。
研究结果表明,NSUN2介导的m5C RNA甲基化在RB的致癌过程中促进嘌呤的生物合成。
- 与正常视网膜相比,RBs中的全局和mRNA m5C水平显著富集。
- 在RB样品和细胞系中NSUN2的肿瘤特异性表达增加。
- 在治疗上,NSUN2的靶向校正在体外和体内均对RB中表现出有效的治疗功效。
- 通过多组学分析,鉴定出嘌呤生物合成中的重要磷酸核糖甲酰甘氨酸脒合酶(PFAS)是NSUN2的下游候选靶标。PFAS的重新引入在很大程度上逆转了NSUN2缺陷型RB细胞的抑制表型,表明PFAS是NSUN2的功能性下游靶标。
- m5C reader蛋白ALYREF负责识别PFAS的m5C修饰,通过增强其RNA稳定性来增加其表达。
总之,本研究证明了NSUN2对于RB中的致癌基因激活是必需的,从而扩展了目前对肿瘤进展过程中动态m5C功能的理解。由于NSUN2/ALYREF/m5C PFAS致癌级联是重要的RB触发因素,因此本研究表明,靶向m5C重编程治疗策略可能是一种新颖有效的抗肿瘤治疗方法。
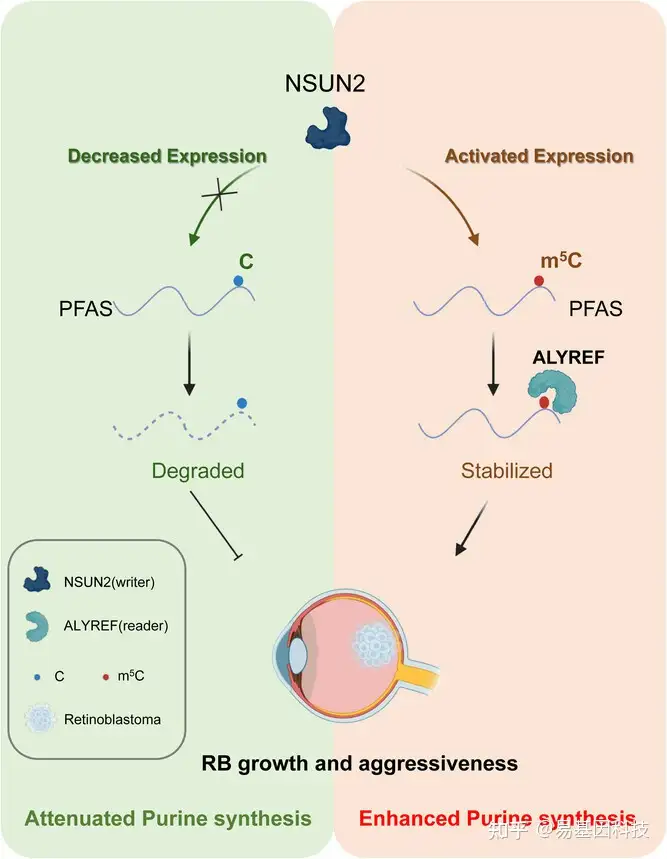
研究示意图:NSUN2在视网膜母细胞瘤细胞中上调增强了m5C甲基化,导致PFAS mRNA和蛋白质表达增加以及腺苷酸(AMP)和鸟苷酸(GMP)含量提高,这些变化促进与视网膜母细胞瘤进展相关的侵袭性增加。
研究结果
(1)NSUN2表达增加增强了RB的m5C修饰水平
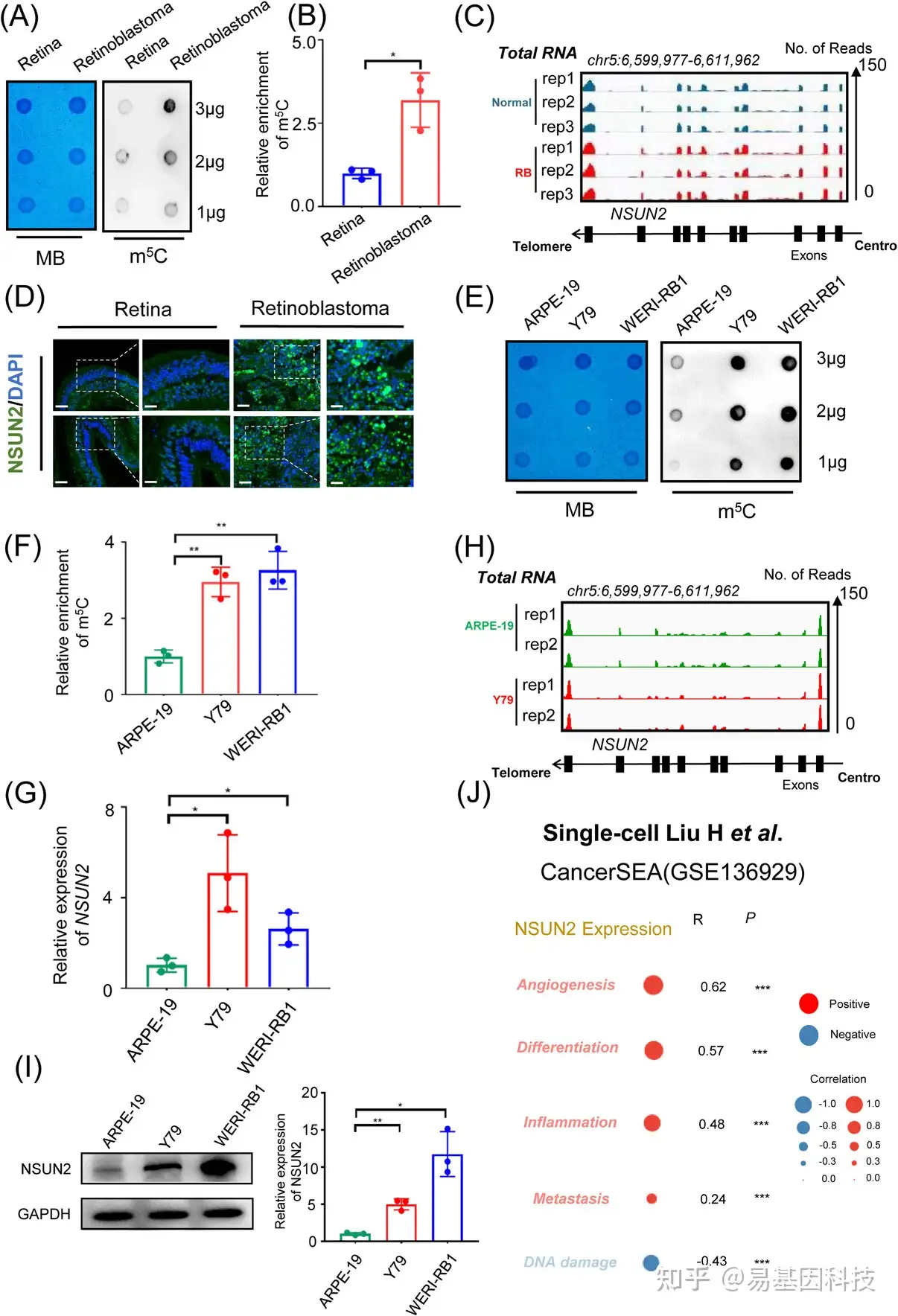
图1:增加的NSUN2表达增强了视网膜母细胞瘤中的m5C修饰水平。
(A-B) 斑点印迹(Dot blot)显示m5C信号与视网膜母细胞瘤和正常视网膜组织中的亚甲基蓝信号的比较。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定*p<0.05。
(C) 整合基因组学浏览器(IGV)显示NSUN2在视网膜母细胞瘤组织中的表达高于正常视网膜组织。
(D) 肿瘤和正常样品中NSUN2(绿色)和DAPI染色(蓝色)的免疫荧光。比例尺:左图,50μm;右图,25μm。
(E-F) 斑点印迹显示与正常视网膜色素上皮细胞系相比,视网膜母细胞瘤细胞系中的m5C信号与亚甲基蓝信号。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定**p<0.01。
(G) qPCR数据显示NSUN2在视网膜母细胞瘤细胞中相对于ARPE-19细胞的表达。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定*p<0.05。
(H) IGV显示与视网膜色素上皮细胞系相比,视网膜母细胞瘤细胞系中NSUN2的更高表达。
(I) Western blot数据和统计分析显示,相对于ARPE-19细胞,视网膜母细胞瘤细胞中NSUN2表达。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定。*p<0.05,**p<0.01。
(J) 单细胞转录组分析揭示了视网膜母细胞瘤中相对NSUN2蛋白表达与不同生物学过程之间的相关性*** p<0.001
(2)NSUN2在体外和体内促进RB的恶性增殖
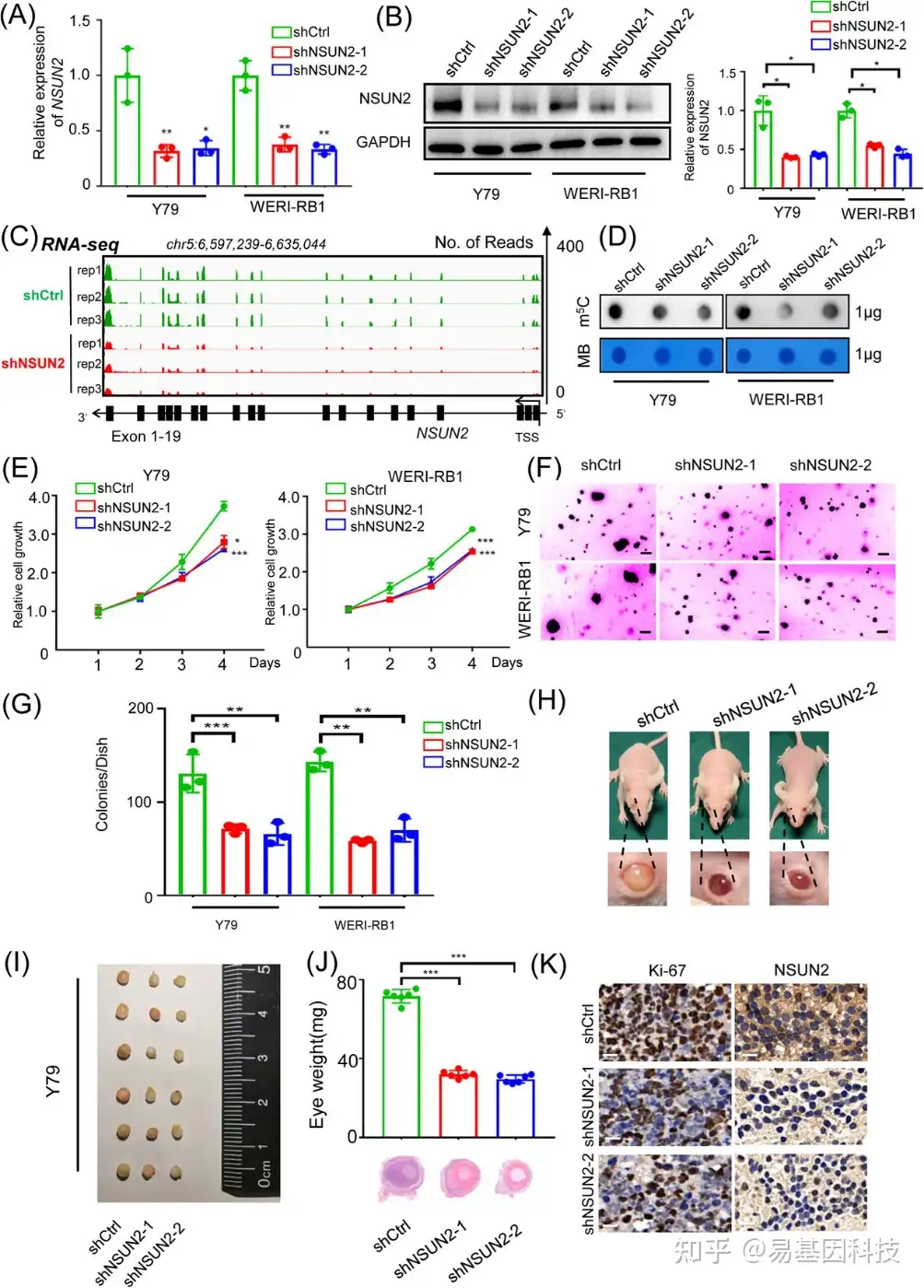
图2:NSUN2在体外和体内促进视网膜母细胞瘤的恶性增殖。
(A) qPCR数据显示NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)中NSUN2表达。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定。*p<0.05,**p<0.01。
(B) Western Blot显示NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)中NSUN2表达。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定。*p<0.05。
(C) IGV显示NSUN2敲低后视网膜母细胞瘤细胞(Y79)中NSUN2表达。
(D) 斑点印迹显示NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)中相对于亚甲蓝信号的m5C信号。
(E) CCK-8实验评估NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)的增殖。数据为三个生物学重复的平均值±SD。显著性由非配对双尾学生t检验确定。*p<0.05,***p<0.001。
(F-G) 采用软琼脂测定法评估NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)的增殖。显示了来自三个重复的代表性图像。数据表示为平均值±SD。显著性由不成对的两尾学生t检验确定**p<0.01,***p<0.001。
(H-I) BALB/c裸鼠和含有源自NSUN2缺陷型Y79细胞的异种移植物的眼球图像。显示六个生物学重复的代表性图像。
(J) 眼球重量数据的统计分析。H&E染色以评估肿瘤形成。数据表示为平均值±SD。显著性由不成对的两尾学生t检验确定***p<0.001。
(K) 免疫组化染色图像显示对照组和NSUN2敲低组中Ki-67和NSUN2表达。比例尺:50μm。
(3)PFAS调控NSUN2的m5C修饰
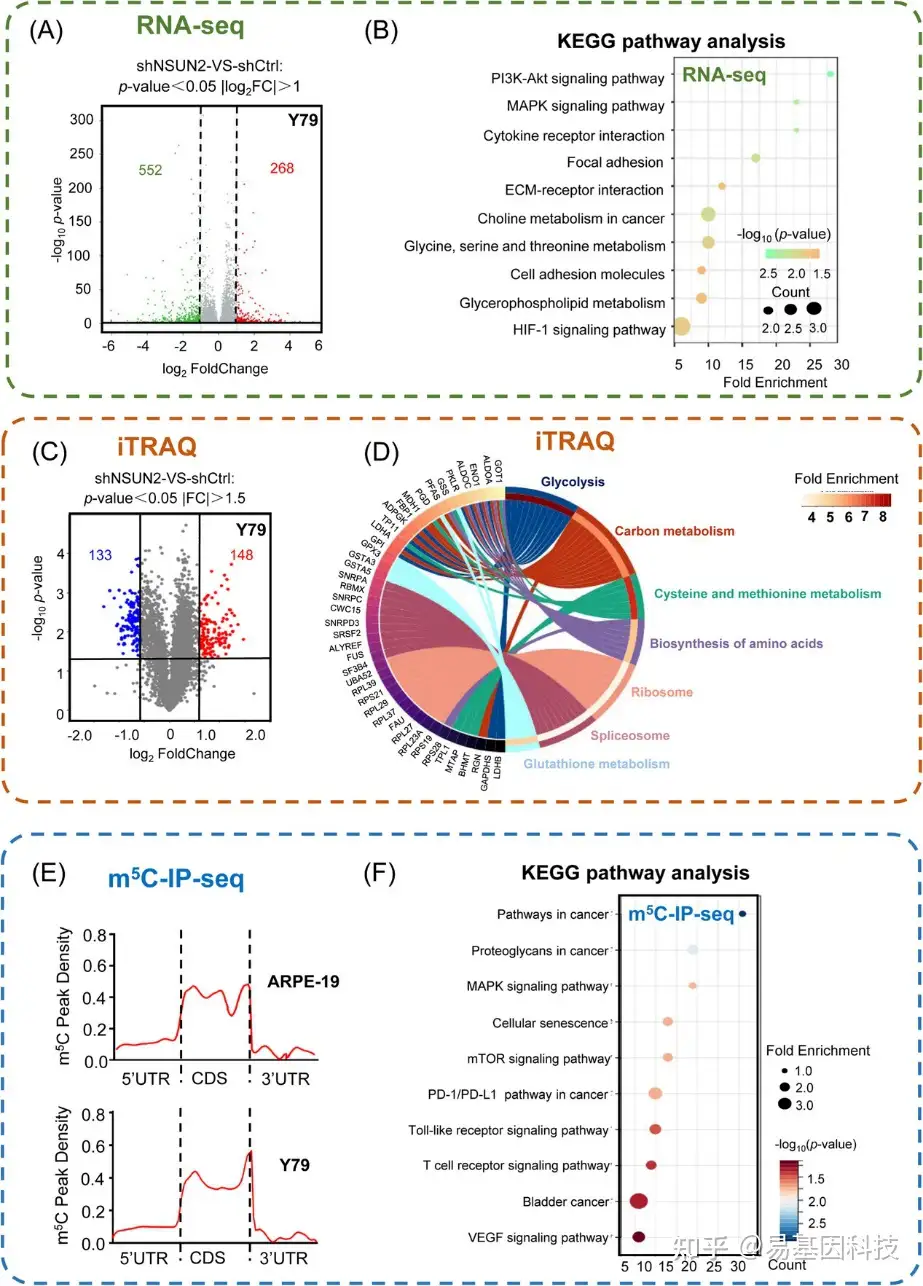
图3:多组学分析以鉴定NSUN2的潜在RNA靶标。
(A) 火山图显示NSUN2缺陷型视网膜母细胞瘤(Y79)细胞中NSUN2调控的基因。
(B) NSUN2缺陷型视网膜母细胞瘤(Y79)细胞中NSUN2调控基因的KEGG通路分析。
(C) 火山图显示NSUN2缺陷型视网膜母细胞瘤(Y79)细胞中NSUN2调控的蛋白质。
(D) NSUN2缺陷型视网膜母细胞瘤(Y79)细胞中NSUN2调控蛋白的GO富集分析。
(E) m5C meRIP-seq数据显示m5C位点的peaks值密度。分析了生物学重复样品。
(F) 视网膜母细胞瘤(Y79)细胞和正常视网膜组织中m5C修饰基因的KEGG通路分析。
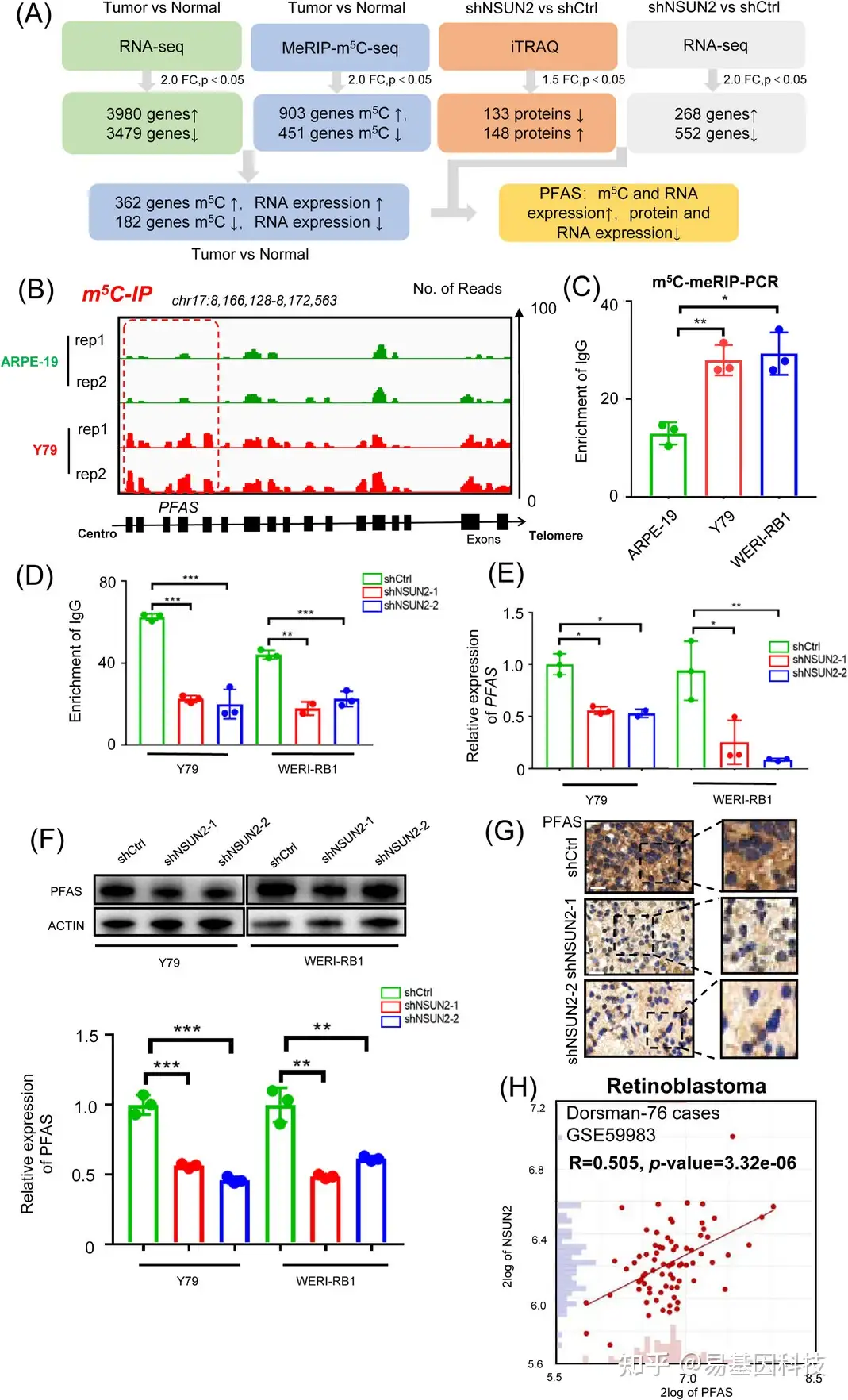
图4:PFAS调节NSUN2的m5C修饰。
(A) 多组学分析将PFAS鉴定为NSUN2的下游标靶。
(B) m5C meRIP-seq分析的IGV轨迹显示PFAS的m5C富集。分析了生物学重复样本。
(C) 视网膜母细胞瘤细胞和视网膜色素上皮细胞中PFAS中m5C状态的m5C-MeRIP qPCR检测。数据三个生物学重复是平均值±SD。显著性由非配对双尾学生t检验确定。*p<0.05,**p<0.01。
(D) m5C-MeRIP qPCR检测NSUN2缺陷型视网膜母细胞瘤细胞中PFAS中m5C状态。数据三个生物学重复是平均值±SD。显著性由非配对双尾学生t检验确定。
(E) qPCR数据显示NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)中PFAS的表达。数据三个生物学重复是平均值±SD。显著性由非配对双尾学生t检验确定。*p<0.05,**p<0.01。
(F) Western Blot分析显示NSUN2敲低后视网膜母细胞瘤细胞(Y79和WERI-RB1)中PFAS的表达。数据三个生物学重复是平均值±SD。显著性由非配对双尾学生t检验确定。**p<0.01,***p<0.001。
(G) 免疫组织化学染色图像显示对照组和NSUN2敲低组中PFAS表达。比例尺:左图,50μm;右图,25μm。
(H) 视网膜母细胞瘤样本队列中NSUN2表达和PFAS表达的相关性分析(n=76)。通过Pearson相关分析确定显著性(R=0.505,p=3.32e-06)。
(4)PFAS是NSUN2的功能性下游标靶
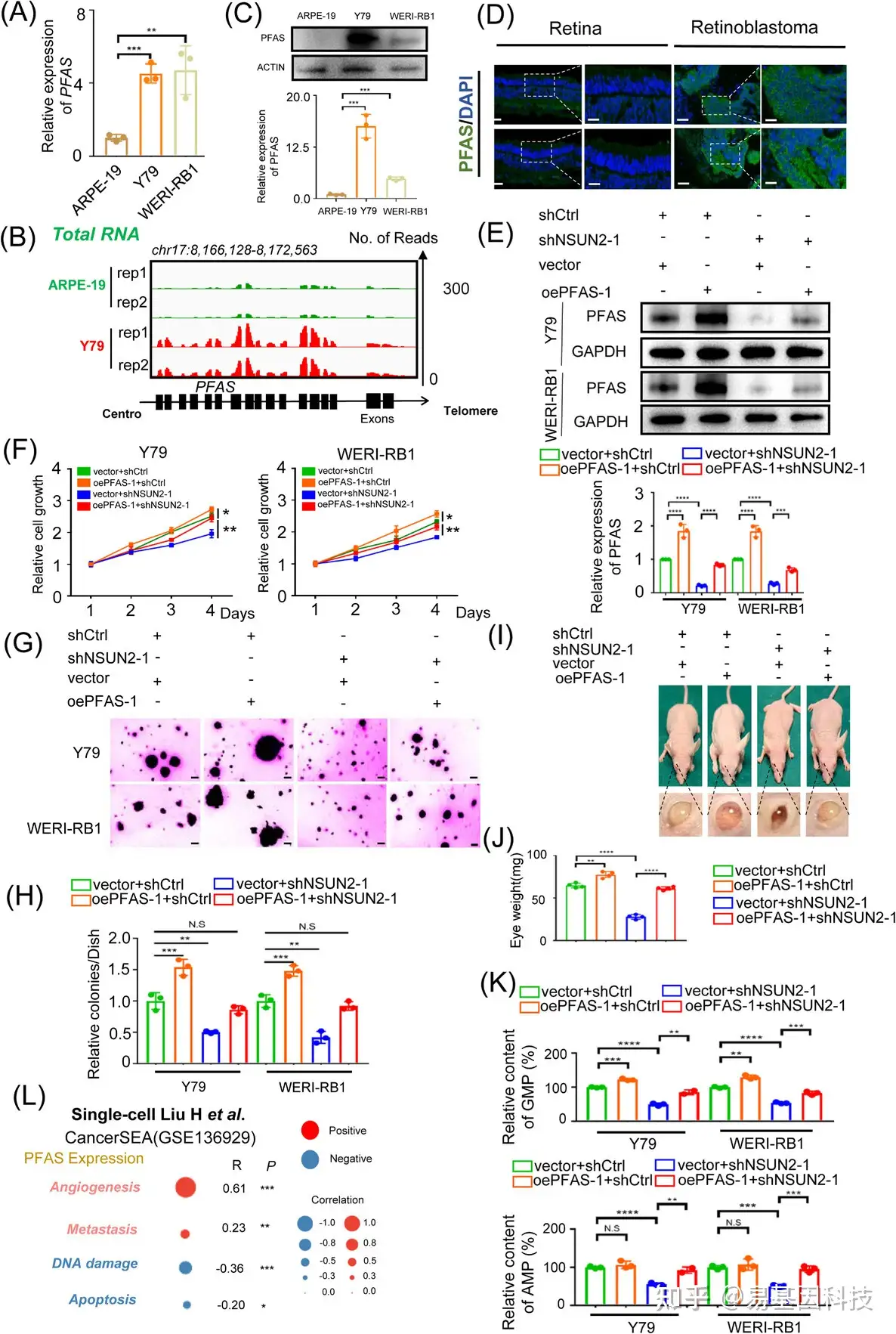
图5:PFAS是NSUN2的功能性下游靶标。
(A) qPCR数据显示与ARPE-19细胞相比,视网膜母细胞瘤细胞中PFAS的表达。数据以三次重复的平均值±标准差(SD)呈现。显著性通过非配对双尾学生t检验确定。**p < .01,***p < .001。
(B)IGV显示与正常视网膜组织相比,视网膜母细胞瘤中PFAS的更高表达。
(C)Western Blot分析显示与ARPE-19细胞相比,视网膜母细胞瘤细胞中PFAS的表达。数据代表三次重复的平均值±标准差。显著性通过非配对双尾学生t检验确定。***p < .001。
(D)免疫荧光显示肿瘤和正常样本中PFAS(绿色)和DAPI染色(蓝色)。比例尺:左图,50μm;右图,25μm。
(E) Western Blot分析显示不同组别视网膜母细胞瘤细胞中PFAS的表达。数据以三次重复的平均值±标准差呈现。显著性通过非配对双尾学生t检验确定。***p < .001,****p < .0001。
(F) CCK8实验评估NSUN2缺陷的视网膜母细胞瘤细胞在PFAS过表达后的增殖能力。数据以三次重复的平均值±标准差呈现。显著性通过非配对双尾学生t检验确定。*p < .05,**p < .01。
(G-H) 软琼脂实验评估NSUN2缺陷的视网膜母细胞瘤细胞在PFAS过表达后的增殖能力。数据以三次重复的平均值±标准差呈现。显著性通过非配对双尾学生t检验确定。**p < .01,***p < .001。N.S.表示无显著性。
(I-J) BALB/c裸鼠和含有源自NSUN2缺陷且PFAS过表达的Y79细胞的异种移植物的眼球图像。眼球重量数据的统计分析。数据以平均值±标准差呈现。显著性通过非配对双尾学生t检验确定。**p < .01,****p < .0001。
(K) 在NSUN2缺陷的视网膜母细胞瘤细胞中检测到PFAS过表达后的腺苷酸(AMP)和鸟苷酸(GMP)浓度。
(L) 单细胞转录组分析揭示了在视网膜母细胞瘤中相对PFAS蛋白表达与不同生物过程之间的相关性。**p < .01,***p < .001。
(5)ALYREF识别m5C甲基化的PFAS并促进其RNA稳定性
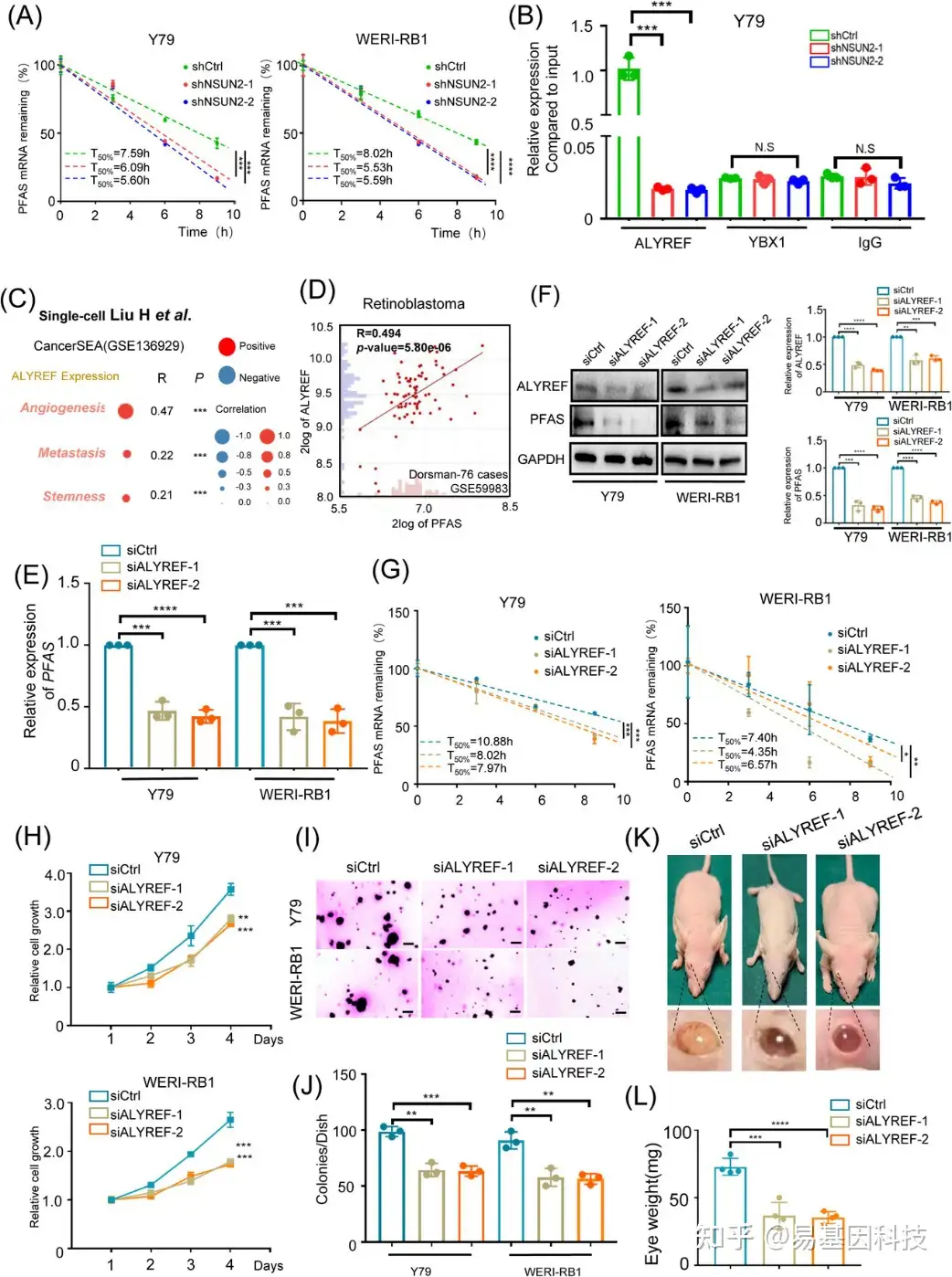
图6:ALYREF识别m5C甲基化的PFAS并促进其RNA稳定性。
(A)在用放线菌素D(10 μg/μL)处理0-10小时的NSUN2缺陷的视网膜母细胞瘤细胞中PFAS的半衰期。
(B)RIP-qPCR检测ALYREF和YBX1诱导的视网膜母细胞瘤细胞(Y79)中PFAS的表达。
(C)单细胞转录组分析揭示视网膜母细胞瘤中相对ALYREF蛋白表达与不同生物过程之间的相关性。
(D)视网膜母细胞瘤样本队列中ALYREF表达和PFAS表达的相关性分析(n=76)。
(E)qPCR数据显示ALYREF敲除后视网膜母细胞瘤细胞(Y79和WERI-RB1)中PFAS的表达。
(F)Western Blot分析显示ALYREF敲除后视网膜母细胞瘤细胞(Y79和WERI-RB1)中PFAS的表达。
(G)放线菌素D(10 mg/mL)处理ALYREF缺陷型视网膜母细胞瘤细胞0-10小时PFAS的半衰期。
(H)CCK-8实验评估ALYREF敲除后视网膜母细胞瘤细胞(Y79和WERI-RB1)的增殖能力。
(I-J) 软琼脂实验评估ALYREF敲除后视网膜母细胞瘤细胞(Y79和WERI-RB1)的增殖能力。
(K-L) BALB/c裸鼠和含有源自ALYREF缺陷的Y79细胞的异种移植物的眼球图像。眼球重量数据的统计分析。
研究小结:
这项研究最初证明了NSUN2通过增强其在RB中的RNA稳定性来激活PFAS,扩展了目前对肿瘤进展过程中动态m5C功能的理解。值得注意的是,RNA上的m5C修饰对于嘌呤合成很重要,弥合了目前对RNA修饰和代谢重编程的理解。由于NSUN2/ALYREF/m5C PFAS致癌级联是RB的重要触发因素,因此本研究提供了一种新的治疗策略,即“靶向m5C重编程策略”,这可能是一种有效的抗肿瘤治疗方法。
关于易基因RNA m5C甲基化测序(RNA-BS)技术
m5C是RNA百余种修饰中研究较多的一种。m5C存在于tRNA上时,可以对翻译进行调节;存在于rRNA上时,可以对核糖体的生物合成进行质控;存在于mRNA上时,则可以影响mRNA的结构、稳定性及翻译过程。
易基因提供适用于不同科研需求的m5C甲基化测序技术:
- 常规mRNA m5C甲基化测序(RNA-BS):
mRNA分离后首先通过亚硫酸盐处理,非甲基化的C转变为U,m5C修饰的碱基保持不变,结合高通量测序,可以对RNA上的每一个C碱基修饰进行定位与定量。 - 常规mRNA +lncRNA m5C甲基化测序(全转录组RNA-BS):
易基因科技建立的升级版m5C RNA甲基化测序服务,去除人rRNA后,剩余RNA经重亚硫酸盐处理后,结合高通量NGS策略,可在全转录组范围内单碱基分辨率地检测基因m5C甲基化修饰分布。
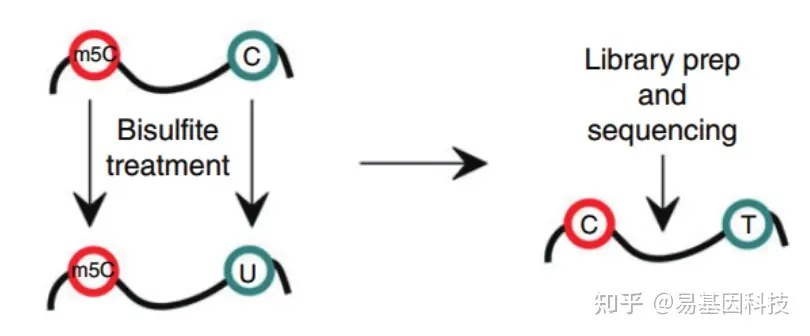
技术优势:
- 高深度:超高深度重亚硫酸盐处理,检测准确性极高;
- 高通量:结合高通量NGS,全转录组范围内检测;
- 单碱基:单碱基分辨率,快速检测和分析RNA中的m5C。
- 高准确:精确的检测mRNA等每一个C碱基的的修饰水平。
研究方向:
- 与m6A甲基化类似,m5C甲基化已被证明与肿瘤、神经系统紊乱、代谢性疾病、病毒感染以及个体发育等密切相关。
- 此外,RNA甲基化(m5C)与人类疾病密切相关,其功能涉及调控干细胞应激、细胞毒性应激、mRNA出核和植物细胞发育及基因表达等方面。
实验策略:
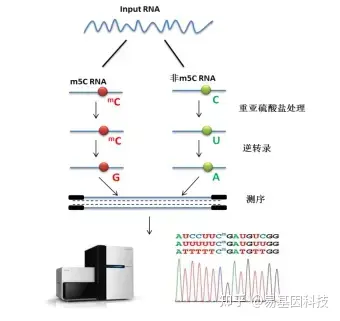
易基因RNA m5C甲基化建库测序示意图
易基因提供全面的表观基因组和表观转录组研究解决方案,详询易基因:0755-28317900.

参考文献:
Zuo S, Li L, Wen X, Gu X, Zhuang A, Li R, Ye F, Ge S, Fan X, Fan J, Chai P, Lu L. NSUN2-mediated m5 C RNA methylation dictates retinoblastoma progression through promoting PFAS mRNA stability and expression. Clin Transl Med. 2023 May;13(5):e1273. doi: 10.1002/ctm2.1273. PubMed PMID: 37228185.
相关阅读:
项目文章 | NAR:RCMS编辑系统在特定细胞RNA位点的靶向m5C甲基化和去甲基化研究
干货:手把手教你做RNA m5C甲基化测序分析(RNA-BS)
标签:视网膜,RNA,NSUN2,甲基化,PFAS,m5C,母细胞 From: https://www.cnblogs.com/E-GENE/p/18101016